-
My archives
pétrole-prébiotique 2011 évolution de la membrane prébiotique 2014 Poster 2014 chiralité-prébiotique 2012 Historique des écrits du 14.3.14 pour les séminaires 2014
-
Par mekali le 19 Juillet 2014 à 12:12
13.7.14 Kyoto
Le speach de Pohorille m'a suggéré qu'il faut considérer la perméabilité des liposomes non pas d'un liposome parfait, mais d'un liposome formé à partir de la soupe prébiotique contenant dans la bicouche de nombreuses molécules qui la rendent entièrement une passoire et qu'elle évolue vers le liposome parfait en gardant juste quelques canaux et pendant cette évolution le métabolisme peut s'initialiser et évoluer, la ségrégation K+/Na+ peut se faire, le phosphate peut entrer dans la phase de séquestration et sa pénétration assurée par des canaux qui auront évolués toujours dans le sens inverse de l'évolution à partir d'un liposome parfait.
Le moteur de cette évolution est l'établissement du potentiel chimio-électrique et la ségrégation Na+/K+ conséquente. D'où la nécessité chez les microbes actuels d'avoir une paroi pour filtrer et un périplasme étroit qui contrôle les protons H+.
15.7.14 Nara suite
Le passage de vésicule à liposome est d'une telle force ( comme les forces de surface qu'on arrive à concrétiser, conceptualiser, pour les insectes qui marchent sur l'eau, ou bien la sortie des larves de l'eau vers l'air) qu'il y a un remaniement des forces mises en place dans la phase huile.
Une fois la bicouche formée la cohésion mécanique réalisée par les acides aminés de configuration L, la renforce de plus en plus. Donc ce passage est la clé de l'évolution de la bicouche lipidique vers le liposome qu'on connaît avec ses protéines ( canaux et complexes énergétiques ) et de l'initialisation du métabolisme. Cette évolution ( potentiel électrique et cohésion ) est une contrainte pour la mise en place du métabolisme.
S'établit alors une triple synergie: cohésion, évolution, métabolisme. Métabolisme qui fournit les acides aminés pour la cohésion et pour les canaux d'échange.
L'évolution de la membrane se dirige vers un potentiel chimio-électrique de plus en plus fort et en résonance avec les actions du milieu extérieur. Toutes les propriétés des 5 zones décrites dans chimio-osmose prébiotique, sont en action. Ces propriétés ( graduation de l'hydropathie ) sont exaspérées au moment du passage vésicule-bicouche.
Le seul point qui reste c'est la synthèse des acides nucléiques. L'évolution par morceaux des acides nucléiques peut commencer avec celle des mono-nucléotides car induite par l'évolution de la bicouche qui a besoin de plus en plus d'acides aminés pour la catalyse ( ATP ), la cohésion ( CTP ) et la mise en place de la paroi ( UTP ). Le GTP ressemble à l'ATP.
L'évolution du métabolisme va introduire les autres coenzymes qu'on a vu dans chiralité notamment B6, NAD, FAD....
Par contre la synthèse des oligo puis des polynucléotides parait inexistante. Pour qu'elle puisse être initialisée il faut mettre côte à côte plusieurs ANs et établir la liaison phospho-diester par une ligase. Dans le monde ARN je crois, ( à confirmer ) que c'est l'activité qui est mise en avant pour son évolution. La ligation apparaîtrait en 1er.
Pour ce qui est de mettre côte à côte les ANs, une remarque de Baigl Damien "la membrane chaperonne l'ADN" laisse penser que c'est la membrane. Et ce d'autant plus que mon hypothèse naissante que les ANs se forment quand les bases ( hydrophobes ) présentent leur N (azote) actif vers le "cytoplasme" et que la D-glycéraldéhyde-P y est fixée, me paraît de plus en plus convaincante. Dans cette hypothèse les dRNs se formeraient en 1er car la D-GA-P s'accumule et que l'acétaldéhyde vient de FT et non de formose. La formation des RNs amènerait la fonction de duplication plus facile à réaliser qu'avec les dRNs.
L'interaction du milieu extérieur ( par rapport au liposome ) / ADN se ferait par les " facteurs de transcription" eux-mêmes résultat de l'interaction du liposome avec ce milieu extérieur. Ces facteurs ouvrent l'ADN double-brin et des morceaux d'ARN peuvent se former. Nous avons là le début de la transcription et de la réplication (Okazaki).
Ainsi la ligation ( par groupe d'acides aminés et/ou d'ARN ) est le point central de l'évolution des ANs.
Nous voyons que l'évolution par morceaux inclut maintenant outre les ARNs que j'avais mentionnés, l'ADN prébiotique et la membrane prébiotique.
A noter :
- Directionnalité de l'évolution grâce à la force créée par ce traumatisme.
- La ligation reste à élucider.
- Le feuillet externe peut contenir beaucoup plus de molécules hydrophobes élaborées. Élaborées par une multitude de vésicules avortées.
- La fragilité des liposomes formés (Baigl): d'où nécessite de durée plus longue dans l'huile (baisser la température chez Baigl). A cela il faut ajouter les hautes pressions.
- La ségrégation K/Na peut désormais se faire, mais il faut l'élucider.
- La thymine est à mettre au même niveau que les autres bases dans la soupe: c'est une autre conformation.
- L'ADN va enregistrer les 1ères étapes de l'évolution du métabolisme, de la membrane et de l'ADN elle-même.
21.7.14 Paris
1. Lecture des articles d'Hiroshi Murakami
avril 2011: Determination of Structural Parameters of Protein-Containing Reverse Micellar Solution by Near-Infrared Absorption Spectroscopy. Hiroshi Murakami,Takaki Nishi, and Yuji Toyota.
dx.doi.org/10.1021/jp111852s | J.Phys.Chem.B 2011, 115, 5877-5885
- La myoglobine n'est pas au centre, mais à la surface interne de la micelle. rapport eau/micelle= w0= 2-8.
- Conformation distordue par rapport à la solution aqueuse due à l'interaction avec la tête hydrophile.
- Table 2: eau entre 5 000 et 10 000 molécules; têtes entre 300 et 400; Rayon entre 3-4 nm: spectro proche infra-rouge.
novembre 2011: Terahertz absorption spectroscopy of protein-containing reverse micellar solution.
H. Murakami, Y. Toyota, T. Nishi, S. Nashima
Chemical Physics Letters 519-520 (2012) 105–109
- TeraH w0= 0.2, 4.4
- Relation entre low-frequence motion des protéines et leur fonction biologique ( autour du teraH ). On pense que ce mouvement joue un rôle important dans la fonction des protéines.
- TeraH intéressant dans solvant huile, car l'eau donne un grand bruit de fond.
- nanometer-confinement effect
- L'activité enzymatique est fonction de w0.
- Structure des micelles avec myoglobine dérivée de l'article du 4.11 précédent.
- Révélation de l'importance de l'hydratation des protéines. Les têtes hydrophiles réduiraient les couches d'hydratation.
- Le teraH serait intéressant pour l'étude dynamique de l'activité enzymatique et le repliement des protéines.
Novembre 2013: Nanometer-scale water droplet free from the constraint of reverse micelles at low temperatures.
H. Murakami,T. Sada, M. Yamada and M. Harada
PHYSICAL REVIEW E 88, 052304 (2013) DOI: 10.1103/PhysRpressionsevE.88.052304
- w0<5
- sonde photosensible: rhodamine 6G, très grosse (voir wikipedia).
- Intérêt des micelles reverses pour l'étude du water supercooled: organisation des molécules d'eau.
2. Brouillon pour la lettre
- Hautes pressions
- C/L
- organisation
- problème A/R ----> petits liposomes, quantique à travers la membrane (ref en cours).
- L'intérêt des études dans les micelles ou dans les liposomes ----> les 1ères étapes de l'évolution moléculaire:
- avec les mêmes protocoles ----> relation entre membrane et acides aminés au lieu des protéines,
- le plus important, communication ---> d'où Pohorille sur liposome parfait, Baigl et diffusion ----> évolution de la membrane.
22.7.14 Paris
Cher Iroshi Murakami,
J'ai pu lire tes 3 articles. Ton travail m'intéresse beaucoup parce qu'il rejoint une réflexion que j'ai entamée après mon analyse de la cohésion mécanique du liposome. Tu peux voir cette analyse dans le poster " chiralité prébiotique".
Cette réflexion partait d'un problème que me posait la théorie de la chimio-osmose de P. Mitchell (1960). En effet cette théorie traite du potentiel électrochimique qui, dans la physique classique, suppose un champs électro-magnétique agissant à distance qui établit ce potentiel. Or dans les liquides il n'y a pas de vide et tout champs électro-magnétique local est écranté par les molécules voisines avant d'arriver aux molécules éloignées. Mon étude sur la communication à travers la membrane que j'ai publiée dans chimio-osmose prébiotique se trouvait bloquée alors que j'attribuais au potentiel électrochimique le rôle moteur de l'évolution moléculaire [ 2ème partie du poster "pétrole prébiotique"].
Avec les propriétés de la cohésion mécanique j'ai pensé alors qu'une haute pression hydrostatique rigidifierait la membrane et permettrait la communication à travers la membrane par le processus d'action/réaction comme dans les solides, et les charges accumulées sur la face externe provoqueraient par ce processus l'ionisation des molécules internes qui les neutraliseraient. En fait le potentiel électrique ne fait qu'augmenter.
L'idée derrière les hautes pressions hydrostatiques c'est d'accélérer un processus hypothétique qui ferait pénétrait les acides aminés dans la membrane pour enfin constituer des canaux d'échanges. Pourquoi les acides aminés? Parce que les canaux d'échanges sont protéiques et que les acides aminés sont des zwitterions tout comme les têtes hydrophiles aux quelles ils peuvent s'accrocher et pénétrer ainsi dans la membrane.
Cette hypothèse de pénétration des acides aminés dans la membrane est en fait bâtie sur une idée préconçue qui nous paraît logique. Cette logique est la suivante: l'évolution moléculaire, pour obtenir des canaux d'échanges à travers la membrane, doit augmenter la perméabilité des acides aminés. L'à priori est l’existence préalable d'une membrane parfaite comme elle existe chez le vivant mais sans les canaux. Toutes les expériences sont faites dans cette logique et ma réflexion, jusqu'à la prestation orale du Pr A. Pohorille du 13/7, était basée sur cette hypothèse.
*********
Les idées à la base
- dipeptides Pohorille
- gradient hydrophobicité et solubilité sur les faces des vésicules réverses
- fragilité des liposomes formés suivant mon schéma ( Baigl )
- Les forces surfaciques et la rupture du feuillet externe qui se referme sur le feuillet interne
- dissymétrie entre les 2 feuillets par leur constitution provoque de nombreuses zones à dissymétrie électrique entre les 2 feuillets, donc des potentiels, donc des mouvements d'ions et de molécules avec leurs cortèges de H2O d'hydratation d'où la formation de canaux prébiotiques qui sont au début que des voies de circulation et qui seront tapissés d'acides aminés avec l'évolution moléculaire pour donner les canaux ioniques que l'on connaît.
_ L'évolution moléculaire de la membrane prébiotique.
- La circulation est nécessaire pour l'équilibre des charges et elle est permanente sauf pour les liposomes parfaits.
- d'abord il y a initialisation de la fixation du glycérol-P comme dans "chiralité prébiotique", qui produit des têtes hydrophiles servant d'attraction aux acides aminés pour le processus de cohésion mécanique.
- puis il y a la mise en place de la bicouche avec les forces de V. der Walls des chaînes aliphatiques qui se rapprochent.
- La mise en place des dipôles pour annuler les répulsions électroniques entre ceux dont les chaînes sont courtes.
- Action progressive des acides aminés pour la cohésion mécanique. Elle s'intensifiera quand la sérine sera fixée par liaison ester au P. Cette estérification est favorisée par le mouvement d'ensemble et par la surface polyanionique de la face interne.
- A une certaine concentration la dissymétrie électrique provoquée par la fixation de la surface interne uniquement, doit provoquer la décarboxylation de la sérine au lieu de son enlèvement. Ce qui abaisse la dissymétrie électrique tout en maintenant une forte cohésion mécanique de l'ensemble. Cette décarboxylation est aidée par B6 qui agit en cohésion mécanique comme les acides aminés et comme coenzyme pour les groupes d'acides aminés amassés sur la face interne ( voir initialisation du métabolisme).
- La décarboxylation des autres acides aminés peut être alors envisagée. Les inter-conversions entre acides aminés et les oxo-acides est mise en place. C'est la 1ère étape importante de l'évolution du métabolisme après son initialisation.
_ Interprétation de l'évolution moléculaire de la membrane.
- Ce titre veut dire qu'avant on ne voyait pas l'évolution de la membrane mais l'évolution des canaux. Et A. Pohorille a fait un pas de plus en proposant la co-évolution de la membrane et du métabolisme. Mais les expériences qu'il a présentées montrent bien l'évolution des canaux et du métabolisme, mais il n'y a toujours pas d'évolution de la membrane dans son ensemble comme je l'ai dit ci-haut.
- Je vois maintenant l'évolution de la membrane comme une membrane passoire au début et qui évolue vers une membrane semi-perméable avec des canaux fonctionnels divers. Les pores qui permettaient la circulation de flux importants vont se fermer petit à petit grâce au rapprochement des PLDs par les forces de V. der Walls et la cohésion mécanique pour combattre les fluctuations des potentiels électriques et les canaliser.
C'est une évolution plus physique que chimique:- physique: dipôles, v. der Walls, circulation, cohésion...
- Chimique: 4 estérifications, acides gras, phosphate, glycérol et sérine dans un 1er temps puis décarboxylation de la sérine dans un 2ème temps.
- L'évolution moléculaire de la membrane devient réellement une co-évolution du métabolisme et de la membrane. Deux problèmes qui ne se posent plus alors, ce sont la mise en place de la ségrégation K/Na que je n'arrivais pas à concevoir dans l'autre hypothèse. Reste le pourquoi de cette ségrégation, mais à mon avis cela a peu d'importance pour l'évolution moléculaire en générale et pour l'origine de la vie. Le 2ème problème crucial qui ne se pose plus est la pénétration du P. Comment peut-on envisager la poursuite de l'évolution moléculaire sans P?
_ Le concept global de l'évolution moléculaire.
- Un 3ème problème que devrait résoudre l'évolution moléculaire de la membrane est le concept d' A/R du concept global. Le concept global que j'étudie dans un article en préparation ( ref. ) est le concept de la continuité entre évolution moléculaire et évolution darwinienne. ----
- Rappel des hautes pressions et des milieux condensés en relation avec les 3 articles de H. Murakami.
- Énonce du concept global
- C/L
- organisation
- A/R ---->
- mobilité ----> canaux
- facteurs de transcription prébiotiques ----> ADN
- Problème réplication / duplication
- évolution par morceaux dont membrane.
*****
23.7.14 Paris
_ Étude des milieux condensés.
Si je vous ai écrit en détail ma réflexion sur l'évolution moléculaire de la membrane prébiotique, c'est pour vous montrer combien est complexe l'étude des milieux condensés. Certes c'est une façon de communiquer mes découvertes, mais aussi j'essaie de vous montrer que le travail d'imagination et de réflexion devient obligatoire quand on aborde une si grande complexité.
Et ce travail ne peut être possible que s'il s'appuie sur des concepts simples confirmés par l'expérimentation comme ceux élaborés par la thermodynamique et la mécanique quantique. C'est pour cela que vos travaux et ceux sur les milieux condensés en général sont importants. Car ils nous permettent d'entrevoir le comportement d'une entité individuelle ( un atome, une molécule) dans une situation très simple qui permet l'expérimentation. Je dis entrevoir et même deviner tant la taille de l'échantillon a une très grande importance.
C'est ainsi dans le 1er article du 4-2011 j'ai relevé une idée intéressante. C'est que la myoglobine ne se situe pas au centre de la micelle mais au contact avec les têtes hydrophiles. Ce qui montre l'importance de la relation membrane/protéine dans les recherches sur les origines de la vie. Par contre la conformation de cette protéine est distordue. Ceci est du à la taille de la micelle. Des statistiques ont été faites sue les molécules de la bactérie E.Coli (ref.) et elles montrent qu'elle est cent mille fois plus grande que les micelles que vous étudiez.
L'étude de l'organisation dans une bactérie est à comparer à la gestion d'un pays de quelques dizaines de millions d'habitants. L'expérimentation ne permet que de prendre un échantillon, une classe de collège d'une trentaine d'élèves; les règles d'organisation sociale y sont, mais ceux de la gestion d'un pays entier n'y sont pas. Peut-être que si on avait toutes les molécules d'une bactérie et leurs propriétés, on puisse un jour avec des lordinateurs puissants, lui appliquer les mêmes règles de gestion que l'on applique à un pays.
Dans le 2ème article du 11-2011, la technique du téraHz permet d'étudier les vibrations, les mouvements, l'hydratation et le repliement des protéines toujours dans les micelles reverses de taille nanométrique. Tous ces processus influent sur l'activité enzymatique. Une question importante que je me suis posée sur l'activité enzymatique des protéines dans la cellule et non en solution aqueuse: pourquoi la réaction d'estérification/hydrolyse, la plus utilisée par la cellule est toujours directionnelle ( dans un seul sens ), alors que la réaction thermodynamique ( en dehors de la cellule ) est réversible? C'est comme si cette dernière n'existait pas dans la cellule et d'ailleurs elle ne devrait pas exister puisqu'il y aurait un réseau parallèle au réseau enzymatique qui le contrarierait dans ses contrôles. C'est comme si les réactants et les produits n'étaient pas en contact avec l'eau. Mais comment alors passeront-ils d'un enzyme à l'autre? Les expérimentations en micelles reverses et peut être la technique téraHz pourraient en donner certainement un aperçu.
Le 3ème article du 11.2013 aborde l'organisation des molécules d'eau dans la micelle, ou water supercooled, sous l'influence des têtes hydrophiles. Ces expérimentations sont très importantes pour la recherche sur l'origine de la vie. Ces résultats veulent dire que la micelle ou le liposome, avec leurs têtes hydrophiles créent un début d'organisation. De point de vue conceptuel c'est énorme, car si on ajoute l'organisation de la bicouche et son évolution comme je l'ai présenté précédemment, le problème de l'initialisation de l'organisation est quasi résolu, et ceci grâce à 4 estérifications de 3 petites molécules: glycérol, phosphate et sérine.
J'espère, cher Iroshi que vous allez continuer vos recherches dans cette voie, car elles paraissent très prometteuses pour la recherche sur les origines de la vie. je souhaiterais pour ma part, et comme je l'ai signalé au début de cette note, introduire dans ces expérimentations les hautes pressions hydrostatiques, comme elles sont pratiquées en chimie verte, de quelques kilobars.
Merci encore de vous intéressez à mes travaux.
Berrada Ali Paris 25.7.14
24.7.14 Paris
*******
essais de réécriture de la suite à partir du chapitre "les idées à la base" du 22.7.14
_ 1er essai:
Genèse du scénario de l'évolution de la membrane prébiotique
Je dis bien du scénario et non d'un scénario, parce que j'estime, comme je l'ai montré précédemment, que les expériences faites jusqu'à maintenant concernaient la mise en place de canaux comme si les PLDs étaient inertes, que la membrane n'était qu'une frontière, un contenant.
[ gradient de solvatation de l'interface eau/huile des vésicules aqueuses dans l'huile ( ou des 2 phases principales ) mais pas à travers la bicouche car la phase huile ( queues aliphatiques ) est très limitée. Sous-chapitre "les vésicules de la phase huile" de 5.1.2 de chiralité prébiotique].
_ 2ème essai:
Genèse du scénario de l'évolution de la membrane prébiotique
En entendant A. Pohorille (ref.) dire que les petits peptides accroissent la perméabilité du liposome j'ai eu une réaction étonnante. Je m'entendais dire "mais les liposomes peuvent être perméables à diverses sortes de molécules". En fait je venais de repenser au scénario de la formation des liposomes que j'ai décrit dans l'article "chiralité prébiotique" ( ref au 1er essai ci-dessus). Ce scénario stipule que l'interface eau/huile des vésicules aqueuses dans la poche de pétrole prébiotique constitue un gradient de solvabilité occupé par les molécules de la soupe prébiotique. Mais cette grande perméabilité potentielle ne provoque pas nécessairement la formation de canaux d'échanges.
C'est certainement en poursuivant cette réflexion de membrane passoire et en pensant à la discussion que j'ai eue la veille avec Baigl sur la fragilité des liposomes formés par le processus analogue à celui que je propose dans l'article pétrole prébiotique, qu'il m'est apparu évident que mon scénario de la formation des liposomes était à l'origine de l'évolution de la membrane prébiotique.
******
écrit définitif
Scénario de l'évolution de la membrane prébiotique
Avec ce séminaire, en ayant vu toutes les tentatives pour former les liposomes ou bien pour les remplir de molécules pharmaceutiques, une façon nouvelle m'est apparue pour concevoir la perméabilité prébiotique des liposomes. L'idée serait de partir d'un liposome entièrement perméable ( ou avec de nombreux pores ) qui évoluerait vers un liposome avec un nombre restreint de canaux protéiques comme les liposomes actuels. Ce liposome je l'appelle liposome prébiotique.
_ Formation du liposome prébiotique ( avec des pores ):
Le processus pour former ce liposome est décrit dans mon poster "pétrole prébiotique", et par 2 posters qui travaillent sur l'encapsulation des produits pharmaceutiques ( Baigl et un autre, à voir ). Ce processus est décrit schématiquement et n'aborde pas en détail le rapport des forces en présence.
- Dans la vésicule aqueuse de la phase huile les forces en présence dans le futur feuillet interne sont celles de V. der Walls qui unissent les queues aliphatiques et celles, ioniques, qui rapprochent les têtes hydrophiles zwitterioniques ( cohésion mécanique ).
- Dans l'interface huile/eau principale, futur feuillet externe, les forces sont identiques que précédemment.
- Quand la vésicule aqueuse se rapproche de l'interface principale, 2 types de forces entrent en jeu:
- les forces surfaciques qui font intervenir la résistance à la déformation des l'ensemble des constituants des 2 interfaces. Nous savons en physique qu'elles sont énormes.
- localement des potentiels électriques dus à la différence de constitution des 2 feuillets. Ces potentiels sont toujours présents car le milieu extérieur est changeant par définition.
- Pour qu'une bicouche se forme il faut qu'elle vainc les forces surfaciques et qu'elle annule les potentiels électriques.
- Pour vaincre les forces surfaciques il suffit que la résultante de cohésion du feuillet externe leur soit supérieure.
- Pour annuler les potentiels électriques il faut qu'il y ait circulation des ions, avec leur cortège de H2O d'hydratation, à travers la bicouche. Ce qui n'est possible que si les 2 feuillets étaient de constitution hétérogène. Ce qui est le cas de la soupe prébiotique. Dans le cas des expériences pharmaceutiques il serait difficile d'obtenir des liposomes si on utilisait un phospholipide pur.
La formation des pores sera d'autant plus facile que les feuillets contiennent des phospholipides à courte queue aliphatique. La cohésion mécanique entre phospholipides fait qu'ils se rassemblent en zones à queues de même taille sensiblement. Les zones à queues les plus courtes des 2 feuillets en face l'une de l'autre subissent la répulsion de leurs dipôles et forment des tubes dont les queues aliphatiques pénètrent perpendiculairement dans les zones à queues longues qui constituent la bicouche.
_ Évolution de la membrane prébiotique ( ou bicouche ):
- Les acquis: Avec cette hypothèse du processus de formation des liposomes munis de pores de communication nous avons fait un pas de géant, du moins en théorie. 4 points sont acquis désormais:
- Le nombre de pores prébiotiques par rapport au volume du liposome est limité dans une fourchette étroite déterminée par les forces surfaciques. Les vésicules comportant trop de petites aliphatiques ( donc plus de pores à l'arrivée ) avorteront. Les vésicules comportant trop de queues aliphatiques longues ne pourront pas annuler les potentiels électriques, ne pourront pas accrocher le feuillet externe et resteront dans la phase huile. Cette étape de l'évolution sera très rapide.
- Désormais les potentiels électriques peuvent être annulés de façon régulière en réagissant aux fluctuations du milieu extérieur, et donc jouer leur rôle moteur dans l'évolution moléculaire.
- La ségrégation Na/K peut désormais s'établir sans qu'on ait besoin de savoir pourquoi.
- Le phosphate peut désormais entrer et être séquestré dans le métabolisme naissant.
- Les consolidations à réaliser par l'évolution moléculaire:
Malgré ce pas de géant théorique tout n'est pas acquis cependant et le liposome peut se disloquer parce que très fragile et le métabolisme naissant peut prendre la fuite par les pores. 4 consolidations sont à réaliser par l'évolution moléculaire et dans certains cas rapidement.
- Le nombre de pores peut être trop élevé par rapport au volume et fragiliserait le liposome.
- Le nombre de queues aliphatiques pas assez longues restant dans la membrane la déstabilise par la répulsion des dipôles de leurs têtes.
- Les caractéristiques des pores sont variées et diverses étant issus de la soupe prébiotique, et ne correspondent pas à un fonctionnement cohérent des échanges. Par exemple il se peut qu'il y ait trop de pores à anions ( phosphate ) ce qui empêcherait l'entrée du phosphate; les queues aliphatiques trop courtes pour s'accrocher à la membrane ce qui désorganise le pore et fragilise la membrane.
- La fuite des molécules du métabolisme naissant ralentirait ou stopperait carrément l'évolution moléculaire.
- 2. Le déséquilibre provoqué par les phospholipides à queue aliphatique pas assez longues est une contrainte pour la cohésion mécanique. Nous avons vu dans "chiralité prébiotique" que c'est L-Ser qui joue le rôle principal dans la cohésion mécanique et qu'elle peut être secondée par les autres L-acides aminés quand la tête hydrophile est absente ou incomplète. Cette contrainte à la cohésion mécanique peut accumuler trop d'acides aminés à la surface interne ce qui contraindrait à leur décarboxylation pour maintenir la cohésion mécanique tout en éliminant une accumulation de charges négatives. L'évolution du métabolisme naissant est renforcé puisqu'on passe à une étape ultérieure de son initialisation qu'est l'inter-conversion des acides aminés et des oxo-acides.
- 3. Si on exclut les acides aminés et les hydroxy acides des molécules qui tapissent les pores, celles-ci très diverses et pas assez solidaires entre-elles ne favoriseraient pas l'ordre. Les acides aminés et les hydroxy acides apportent la solidarité entre-eux grâce aux liaisons hydrogènes qu'ils peuvent entre-eux. En plus ils peuvent former ( sans qu'ils soient reliés entre-eux par des liaisons peptidiques ) des hélices alpha qui consolident d'un côté la membrane avec des radicaux aliphatiques et qu'à l'intérieur du pore ils exposent des radicaux ionisés ou polarisés variés qui filtrent de façons très variées et fines diverses molécules ionisées ou polarisées. Les hélices alpha et la membrane se consolident mutuellement.
- 4. La fuite des molécules du métabolisme naissant ne peut être une contrainte, seulement une sélection. Ce sont les métabolismes qui stockent le plus rapidement et le plus efficacement leurs petites molécules qui pourront continuer à évoluer. Par contre l'interaction entre métabolisme et pores est une contrainte pour le 1er. Le métabolisme a besoin de pores efficaces pour interagir avec le milieu extérieur.
- 1. La fragilité entraînée par un nombre de pores excessif est diminuée par l'évolution de la cohésion mécanique. La consolidation des pores par les acides aminés offre par contre une opportunité pour l'évolution de la membrane prébiotique vers les protéines membranaires des systèmes énergétiques en chélatant des métaux de transition nécessaire à ces systmes tels les cytochromes.
 votre commentaire
votre commentaire
-
Par mekali le 2 Avril 2014 à 12:34
Voir préparation du poster 2013 dans continuité-évolution ( à la fin ).
31.3.14 Paris
La continuité entre évolution moléculaire ( E.M : minéral vers procaryote ) et évolution darwinienne ( E.D évolution d'une espèce vivante vers d'autres espèces vivantes ). E.M agit sur des molécules indiscernables et E.D sur des individus discernables.
L'E.D s'effectue sur la complexité. L'E.M s'effectue sur la simplicité, maximale à l'initialisation. L'E.D se manifeste par la constance des caractères ( complexité ) pour l'espèce et leurs variabilité chez l'individu. Dans l'E.M sans complexité on ne peut parler de constance et de variabilité.
2.4.14 Paris
Continuité E.M et E.D: E.M = simplicité, processus séquentiel et parallèle, reproduction du liposome (les 3 articles ); E.D = Complexité, processus à accès direct ( comme en informatique ), réplication de l'ADN mais reproduction de l'ensemble et non de l'ADN seule.
Les 7 principes fondamentaux du concept global: une 1ère approche lors de la réflexion à l'origine de la rédaction des 3 articles.
- Contrainte/liberté: Interface surface minérale/eau produit le pétrole et la soupe prébiotiques, mais aussi les caractéristiques du liposome. Le liposome est une double couche sphérique ( la contrainte ) créant 2 interfaces têtes ioniques/eau avec une double liberté des PLDs et de l'eau.
- Organisation: Auto-assemblage des PLDs en liposome.
- Action/réaction: Ce principe a émergé quand j'ai considéré les hautes pressions hydrostatiques pour le liposome afin que le la chimio-osmose s'effectue plus rapidement et plus efficacement, notamment avec les potentiels protoniques et électroniques.
- Reproduction: C'est la reproduction du liposome. Mais leliposome fait une auto-reproduction à condition qu'on lui fournisse les PLDs; Alors que l'ADN se réplique avec l'assistance des protéines et de l'ARN. C'est une vraie auto-reproduction de l'ensemble puisque la cellule fabrique les propres constituants pour répliquer l'ADN qui lui-même permet la fabrication des protéines et de l'ARN.
- Initialisation: Il faut la soupe prébiotique où tout est possible par déplacement des équilibres chimiques notamment " bases + glycéraldéhyde-P + acétaldéhyde" (ADN), "bases + glycéraldéhyde-P + glycolaldéhyde" (ARN) et "DHA-P + acides gras" ( PLD).
- Production: ou synthèse de nouvelles molécules. Elle se fait avec tous les principes précédents. Pour la reproduction il y a la duplication et l'estérification après appariement.
- Bio-compatibilité: Les conditions nécessaires pour qu'un acide aminé rentre dans la fabrication d'une protéine. La condition suffisante c'est qu'il ne contrevienne pas au fonctionnement d'une protéine qui suit son cycle de sa synthèse et de son hydrolyse: acétylation peptidisation hydrolyse (intéines et hydrolyses, voir famille des aas dans détricotage).
13.4.14 Paris
Rédaction article en vue du résumé pour Poster.
Continuité entre les évolutions moléculaire et darwinienne.
- Introduction: Evol moléculaire. Définition des 2.
- 1ères réflexions (articles) d'après les articles dans l'ordre pétrole chimio chiralité . 4 principes:
- C/L d'après les surfaces minérales puis dans le liposome.
- organisation: auto-assemblage du liposome.
- A/R: haute pression sur liposome pour chiralité et action à distance de la chimio-osmose.
- Reproduction: Celle du liposome mais il faut l'ajout des PLDs.
- 2èmes réflexions (généralisation):
- C/L pour la synthèse des éléments, les cristaux. Contrainte des bases dans l'ADN, liberté dans l'ARN. Les aas piégés par liposome.
- Organisation: éléments organisateurs grâce à leur nuage électronique. Organisation presque cristal dans l'ADN ----> 2 paires de bases. Organisation des aas dans la membrane du liposome.
- A/R: La matière condensée. Les forces dont la gravitation. A/R à travers la membrane du liposome = communication avec l'environnement. A/R ADN facteurs de transcription sans catalyse. La chimio-osmose et la création des canaux ioniques.
- La reproduction: Les éléments sont reproduits par la gravitation. Celle du liposome est triviale à condition d'apporter les PLDs. Celle de l'ADN ne peut être isolée de la reproduction du liposome ou de la cellule.
- Production: Synthèse de liaisons covalentes, amorce les liens nécessaires à la continuité. La synthèse peut se faire dans les principes précédents.
- Initialisation de la continuité avec la soupe prébiotique où existe un réseau de petites molécules déplaçables par les équilibres thermodynamiques. Toutes les molécules de l'initialisation doivent être présentes d'après le principe qu'il n'y a aucun donneur d'ordre. Donc ce sont les réactions demandant peu d'énergie (ester, ions, hydratation ) qui vont fournir les 1ères molécules qui seront piégées, favorisant ainsi le déplacement d'équilibre (formose et non glycérol) = acétaldéhyde, formose, H2 -----> PLDs, ADN, ARN, Thr //aas petits.
- 3ème réflexion(continuité): Délègue ARN pour ADN, et protéines pour liposome. Il n'y a pas reproduction des intermédiaires.
- Transformation des 4 principes, C/L Organisation A/R et Production, du liposome en protéines et ADN en ARN. Les aas procèdent 1 à 1, les nucléiques par segments.
- Il n'y a pas une continuité séquentielle, lipo, prot, ARN, ADN. Les délégations ( et évolution) se font en parallèle. Ce qui fait que l'ADN nécessite 2 paires de bases et pas plus, alors que l'ARN en a une multitude et très catalytique. L'ARN est chélatée par les protéines.
Les protéines + ARN aboutissent au seul et unique code génétique dont le summum de l'évolution se retrouve dans tRNA synthase ----> code à 3 du à la mécanique. - L'A/R ADN/protéines (facteurs de transcription) est mis en place dès le début comme celui de liposome/protéines.
- Apparition d'un nouveau principe: la "bio-compatibilité" avec la famille des aas -----> propriétés des aas codants. Autre que la cristallisation du principe C/L, ici c'est le blocage par des réactions chimiques (ou liaison H). Principe issu des intéines.
- 4ème réflexion (après Darwin): Ces principes s'appliquent ( même bio-compatibilité) à la société et à l'écologie.
- L'espèce parfaite est l'équivalent du cristal d'où mort.
- Le désordre complet c'est les guerres.
- Une C/L nouvelle apparaît à ce niveau = la planète Terre a une limite. Cette limite est-elle infranchissable? Essaimer sur d'autres planètes paraît la solution. Mais l'interaction entre les êtres est vitale et la séparation par les grandes distances la rend nulle. La solution c'est de ne pas s'éloigner, en produisant des vaisseaux géants. Est-ce là la clé du paradoxe de Oppenhimer? Étant donné l'âge de l'univers, de la durée de l'évolution sur Terre, cela ferait longtemps qu'on aurait trouvé d'autres civilisations avancées techniquement pour pouvoir communiquer avec elles.
20.4.14 Tanger
J'ai décidé maintenant de partir des 3 articles dans l'ordre historique de leur écriture pour expliquer l’enchaînement des idées. C'est pour moi une façon de consolider mes résultats. Sinon il y a tellement de points à considérer ( voir livre de Luisi Luigi) que je ne saurais plus pourquoi je prends tel ou tel chemin de réflexion.
- Impasse des systèmes ouverts avec un coenzyme entouré de qq aas (2 ans ).
- La voie des liposomes apparaît très vite prometteuse notamment par la délimitation d'un intérieur où peut progresser l'ordre, et la possibilité d'une synthèse abiotique de leurs acides gras. Seulement cette synthèse se confronte à la polémique du pétrole abiotique.
- Aussi j'ai posé comme postulat de départ pour l'investigation sur les origines de la vie, le postulat suivant:
"La classe des molécules les plus complexes du vivant qui puissent avoir une origine géochimique et abiotique c'est la classe des acides gras à longue chaîne aliphatique". - Pétrole prébiotique.
- À partir de là, comme le pétrole abiotique paraît se former dans la croûte terrestre, j'ai cherché à établir l'état des recherches sur la constitution d'une soupe prébiotique en profondeur de la croûte terrestre et non en surface, car aux acides gras il faut ajouter NPS. Cette idée me paraissait dès le début contre-intuitive pour l'azote et le soufre, car l'azote est essentiellement dans l'atmosphère et le soufre et ses dérivés inhibent la synthèse du pétrole synthétisé par le procédé FTT, utilisé en industrie. Et ce d'autant plus que j'étais à contre-courant des théories et de leurs résultats sur les origines de la vie, théories qui partent de l'expérience de Miller-Urey qui elle se déroulerait dans une atmosphère primitive riche en N2 et qui n'existe plus à cause de l'apparition de l'oxygène O2.
Mais cette soupe prébiotique sous-terraine que j'ai appelée "pétrole prébiotique" a un avantage énorme par rapport à celle issue de l'atmosphère primitive c'est qu'elle est actuelle. Il suffit de démontrer son existence géochimique même indirectement à cause de l'omniprésence du pétrole fossile. Cet avantage est aussi d'ordre conceptuel. Son existence n'est pas due à un phénomène unique du passé, qu'on ne peut plus reproduire, mais qu'elle est due à un processus qui se produit continuellement et donc qu'on peut reproduire nous-mêmes. - Pour compléter la voie de recherche à partir du liposome, dans l'article "pétrole prébiotique" j'ai établi l'état des recherches sur la synthèse des têtes hydrophiles qui nécessitent du glycérol et/ou sérine, éthanolamine, choline. Et j'ai terminé cet article avec la formation des liposomes. L'initialisation du métabolisme écrit à la fin est un résumé des articles "chimio-osmose prébiotique" et "chiralité prébiotique".
- À partir de là, comme le pétrole abiotique paraît se former dans la croûte terrestre, j'ai cherché à établir l'état des recherches sur la constitution d'une soupe prébiotique en profondeur de la croûte terrestre et non en surface, car aux acides gras il faut ajouter NPS. Cette idée me paraissait dès le début contre-intuitive pour l'azote et le soufre, car l'azote est essentiellement dans l'atmosphère et le soufre et ses dérivés inhibent la synthèse du pétrole synthétisé par le procédé FTT, utilisé en industrie. Et ce d'autant plus que j'étais à contre-courant des théories et de leurs résultats sur les origines de la vie, théories qui partent de l'expérience de Miller-Urey qui elle se déroulerait dans une atmosphère primitive riche en N2 et qui n'existe plus à cause de l'apparition de l'oxygène O2.
- La chimio-osmose prébiotique.
Arrivé à ce stade de réflexion, on peut se demander est-ce que la voie du liposome, tel qu'il est actuellement, peut-elle amorcer l'évolution moléculaire? Car si on n'arrive pas à faire communiquer l'intérieur et l'extérieur du liposome, aucune évolution ultérieure n'est possible. Donc, avant de faire des élucubrations sur la synthèse des têtes hydrophiles, j'ai fait une recherche sur la possibilité de créer des canaux d'échange simples à travers le liposome. Or les recherches bibliographiques sur les liposomes aboutissent rapidement sur le processus de chimio-osmose de Peter Mitchell (1961) qui fait intervenir des protéines membranaires et des canaux ioniques eux-mêmes protéiques. Le processus de chimio-osmose est sous-tendu par le potentiel électrique qui se crée entre l'intérieur et l'extérieur.- Il est évident alors que si on arrive à synthétiser un liposome comme l'actuel, mais sans les protéines, ce potentiel existerait aussi. Ce potentiel je l'ai appelé "potentiel électrique prébiotique" et il sous-tend une chimio-osmose prébiotique qui n'a rien à voir en puissance avec la chimio-osmose actuelle.
J'ai émis alors l'hypothèse que cette chimio-osmose prébiotique, même si elle s'avérait très faible, favoriserait l'incorporation des aas dans la bicouche du fait que ces derniers ont la possibilité de s'accrocher à la face externe du liposome, liposome et aas ayant des têtes zwitterioniques de conformation très proches.
L'exemple de canaux d'échange simples, comme je le cherchais auparavant, ce sont les ionophores qui sont des peptides de quelques acides aminés et d'alpha-hydroxydes acides, tous les deux D ou L, et qui permettent des échanges d'ions mono-atomiques dont le proton. - L'article ébauche l'évolution vers d'autres protéines membranaires, mais le plus important ce sont les conséquences conceptuelles sur l'évolution moléculaire:
- Nous avons là un réseau d'électrons et de protons à travers la membrane à la place d'un réseau de réactions chimiques en système ouvert.
- Les cations alcalins sont aussi importants que NPS
- Les toutes 1ères étapes de l'évolution moléculaire accentuent la différenciation entre intérieur et extérieur et notamment par le tri entre des aas vers le feuillet interne.
- La cohésion des feuillets grâce à la structure zwitterionique de la tête et surtout grâce à la longueur du cation ( sérine éthanolamine choline) devrait imposer la conformation des aas et notamment leur chiralité ainsi que celle du glycérol de la tête hydrophile.
- L'ion phosphate doit être traité à part, car il constitue l'armature des feuillets et sa taille et sa charge ne lui permettent pas de passer par diffusion comme les ions mono-atomiques. J'ai suggéré que le processus de flip-flop ( sans enzymes) devrait expliquer son entrée dans le liposome. Son incorporation dans le liposome central et les acides nucléiques constitue ce que j'appelle la séquestration du phosphate.
- Les 5 zones: la membrane n'est pas un contenant seulement (Pohorille).
- L'article chimio-osmose prébiotique laisse en suspens la discrimination entre Na+/K+ et traite le potentiel électrique de façon inadéquate qui est celle du point matériel de la physique classique et notamment les actions à distance entre l'intérieur et l'extérieur comme le suggère la notion de potentiel électrique même. Ces 2 aspects ont été traités après la rédaction de chimio-osmose prébiotique et chiralité prébiotique dans continuité entre EM et ED.
- L'article n'explicite pas clairement la chiralité du glycérol-P, et encore moins l'homochiralité des aas d'autant plus que les ionophores arborent les 2 chiralités de leurs monomères. Une hypothèse que j'ai échafaudée dans l'article conclut que les 2 chiralités du glycérol et de la sérine doivent être complémentaires ( et par extension à tous les aas ). Elle part de la représentation de la tête du phosphatidyl-glycérol où les 2 glycérols sont attachés au même phosphate: glycérol-P-glycérol. L'encombrement stérique étant le même pour les 2 glycérols, leur position par rapport au phosphate rend leur chiralité complémentaire. Mais chez les archées le glycérol (ou la sérine) du cation de la tête hydrophile ne change pas sa chiralité, alors que le glycérol estérifiant les 2 acides gras, oui. L'article chiralité prébiotique traite de ce problème.
- Il est évident alors que si on arrive à synthétiser un liposome comme l'actuel, mais sans les protéines, ce potentiel existerait aussi. Ce potentiel je l'ai appelé "potentiel électrique prébiotique" et il sous-tend une chimio-osmose prébiotique qui n'a rien à voir en puissance avec la chimio-osmose actuelle.
- La chiralité prébiotique.
- La tête hydrophile comporte 2 bras, le fixe celui qui contient le glycérol estérifié à 2 acides gras, et le mobile qui porte un cation (sérine, éthanolamine, choline) ou un glycérol qui sont attirés par l'anion phosphate d'un PLD voisin. Dans cet article je démontre que la chiralité du bras fixe est déterminée par l'encombrement stérique des liaisons esters (L-glycérol-3P) pour les bactéries et celui des liaisons éthers (D-glycérol-3P) pour les archées.
Je démontre aussi que la chiralité du bras mobile portant la sérine ou le glycérol est de type lévogyre (L) rapprochant les 2 feuillets du liposome quand le bras se déplace vers l'anion P, augmentant ainsi la cohésion mécanique du liposome. - Synthèse de la tête hydrophile:
- En prébiotique.
On peut supposer que pour le bras mobile l'estérification pourrait se faire après son positionnement. Je pensais aussi que l'éthanolamine devait être la molécule prébiotique à être estérifiée en 1er étant donné sa simplicité et ne demandant aucun positionnement nécessaire à un carbone chiral.
Par contre pour le bras fixe la difficulté à obtenir du glycérol dans le pétrole prébiotique, le fait qu'il faille créer 2 liaisons esters sur un même glycérol et devant le grand nombre de conformations à traiter si on devait passer par un intermédiaire phosphaté du glycérol m'ont poussé à émettre l'hypothèse suivante: "La DHA phosphatée devrait être hydrogénée et estérifiée sur place, la surface poly-anionique facilitera l'hydrogénation et la tautomérie céto-énol du DHA s'adaptera à 2 acides gras voisins disposés en tête-bêche (se partageant 2 liaisons hydrogènes) pour créer la conformation du glycérol à encombrement stérique minimal. Dans cette hypothèse le L-glycérol-3P, issu d'une éventuelle hydrogénation du glycéraldéhyde-3P sera fixé, alors que le D sera exclu, d'après ma démonstration citée ci-dessus. - En biotique.
- Le bras fixe:
Le D-glycéraldéhyde n'est pas piégé par la membrane et sert l'homochiralité des sucres; Les enzymes qui fixent le glycérol sont membranaires ( c.a.d que la réaction se fait sur la surface), soit elles hydrogènent la DHA-P avant estérification (réaction EC.111.94, réversible comme si elle était calquée sur l'isomérisation DHA-P = L-glycéraldéhyde-3P) soit il y a une 1ère estérification puis hydrogénation (EC 111.54 irréversible) suivie de la 2ème estérification. Les estérifications sont irréversibles. Donc l'hypothèse du DHA ci-dessus semble renforcée.
Cas du L-glycéraldéhyde-3P: Cette molécule n'existe pas apparemment dans le métabolisme. Pourtant une enzyme spécifique de son hydrogénation a été isolée et le L-glycérol-3P qu'elle donne est fixé à 81% dans la membrane (ref.); Les auteurs se posent la question de l'origine de ce glycéraldéhyde. C'est comme si la DHA libre n'existait pas pour la donner par équilibre thermodynamique. - Le bras mobile:
Étonnamment l'éthanolamine que je pensais la molécule la plus simple et la plus facile à installer s'avère indésirable. Chez les procaryotes elle n'existe pas sous forme libre, sinon il existe une seule enzyme pour la casser en NH3 et acétaldéhyde (EC. ). Ensuite l'attachement au phosphate ne se fait pas directement, alors qu'en dehors de la membrane (enzymes non membranaires EC. et EC. ) le phosphate libre et la choline ou l'éthanolamine libres peuvent être liés (chez les eucaryotes seulement).
En fait il y a 2 processus mis en cause dans la fixation du bras mobile. Le 1er est le processus énergétique: la liaison bras fixe- bras mobile est une liaison phospho-diester analogue à celle des acides nucléiques. Alors que les liaisons ester qu'on a vu jusqu'à maintenant sont des liaisons simples. La 2ème liaison est plus énergétique, elle est précédée par une liaison avec un phosphate dans le CDP qui nécessite une 2ème enzyme pour le remplacer. D'où l'intervention du CTP.
Le 2ème processus mis en cause est d'ordre conformationnel. 3 conformations sont à gérer:
- Positionner le cation ( ou le OH pour le glycérol ): C'est lié à l'encombrement stérique du bras. Il faut que le bras soit encombrant (en volume et en charge) pour qu'il puisse se positionner. C'est le cas de la sérine, de la choline et du glycérol-P (différenciation entre L et D). Ce n'est pas le cas ni de l'éthanolamine ni du glycérol. L'éthanolamine est obtenue à partir de la sérine déjà positionnée, par décarboxylation (EC. ), le glycérol est obtenu par déphosphorylation (EC. ) du glycérol-3P installé auparavant (EC. ).
- Positionner la base nucléique nécessaire à la liaison phospho-diester intermédiaire: ce positionnement s'apparente au piégeage des aas avec leur zwitterion. En effet c'est le CDP avec son NH2 et son sucre qui s'apparente le plus à un acide aminé, parmi les bases nucléiques. Les purines et l'uracile ne conviennent pas par leur taille ou par leur manque de NH2 (uracile). Le dCDP joue le même rôle et on a mesuré un rapport de 0.88 entre les acides nucléiques (ref.). Le rôle de positionnement du CDP est nettement séparé de son rôle énergisant (EC. EC. ) où le phosphate du bras fixe n'est plus libre de tourner autour de la liaison phosphoester.
- Positionner le tétraèdre du P du bras fixe qui peut encore tourner autour de sa liaison avec le glycérol. Ce positionnement n'est pas évident mais on le devine avec EC. qui l'élimine. Mais aussi avec le phosphatidyl-CDP où le 2ème P doit bloquer le 1er.
- Le bras fixe:
- En prébiotique.
- Homochiralité prébiotique et initialisation du métabolisme 29.4.14 Tanger.
- Les sucres dérivent du D-glycéraldéhyde-3P qui n'est pas fixé par la membrane. D'où leur homochiralité.
- Pour les aas dans le cadre de la cohésion mécanique, nous n'avons démontré que la chiralité de la sérine. On peut attribuer l'homochiralité des aas à la théorie de la chimio-osmose prébiotique plus la théorie de la cohésion mécanique. Ainsi les mouvements des bras mobiles des têtes hydrophiles entraîneraient les L-aas mais pas les D-aas. C'est ainsi que se formeront dans la membrane des pseudo-hélices alpha de L-aas, d'hélicité droite, qui seront à l'origine des canaux ioniques et des protéines membranaires ( chimio-osmose prébiotique).
Seulement la synthèse des bras mobiles, bien qu'elle nécessite qu'une seule estérification, paraît inaccessible parce que cette estérification, la 2ème d'une liaison phospho-diester, est très énergétique et qu'elle ne bénéficie pas de l'effet de surface comme le bras fixe. Deux processus peuvent accélérer conjointement cette estérification:
- La synthèse des bras fixes crée des surfaces polyanioniques plus ou moins grandes de phosphates ( PO4--, PO4H- ) qui joueront le même rôle que les anions carboxyliques pour le bras fixe, mais beaucoup moins efficacement parce que moins étendues.
- Les L-aas avec leur zwitterion peuvent participer à la cohésion mécanique comme la sérine. C'est ce que j'appelle la séquestration des aas, différente du piégeage des aas par les têtes hydrophiles déjà formées qu'on a vu en chimio-osmose prébiotique. Elle consiste à mettre un L-aa à la place de la sérine quand c'est possible et notamment pour les 1ères mises en place des têtes hydrophiles, dans les espaces sans bras mobile.
- Initialisation du métabolisme.
- Synergie entre cohésion mécanique et L-aas: Le scénario que je décris ici met en oeuvre la synergie entre les L-aas et la cohésion mécanique. Deux cercles vertueux s'établissent alors:
- L'augmentation du nombre de têtes hydrophiles de même chiralité augmente la cohésion de la vésicule, cohésion qui favorise à son tour la fixation et la séquestration des molécules de même chiralité.
- La concentration des L-aas par séquestration crée des groupements d'aas catalysant de plus en plus efficacement la synthèse des têtes hydrophiles mais aussi les 1ères voies métaboliques.
- Le tableau des étapes du scénario (Tableau 1):
- Le scénario commence par la synthèse des têtes hydrophiles dans le cadre de la théorie de la cohésion mécanique ( étapes 1 et 2) et se poursuit en supposant que certains coenzymes puissent être séquestrés comme les 1ers aas (étapes 3 à 7). Ce qui permet aux groupements des L-aas sur la paroi de synthétiser de nouveaux L-aas et nouveaux coenzymes.
- L'initialisation du métabolisme est supposé se produire dans les vésicules aqueuses de la phase huile. Elle peut se poursuivre dans les liposomes dont la membrane n'est pas nécessairement recouverte entièrement de têtes hydrophiles. Nous retrouvons là les processus de la chimio-osmose prébiotique qui vont suppléer en molécules épuisées grâce à la diffusion passive et aux échanges par l'intermédiaire des protéines membranaires.
Dans le liposome prébiotique 2 métabolisme qui se complètent sont mis alors en place:
- Le métabolisme membranaire issu du feuillet externe et qui est sous-tendu par la chimio-osmose qui piège les L-aas externes et les coenzymes minéraux pour constituer les protéines membranaires.
- Le métabolisme de la paroi du feuillet interne en contact avec le milieu intérieur. Ce métabolisme séquestre au début les L-aas et les coenzymes les plus simples sur la paroi. Il emmagasine aussi le phosphate par liaisons covalentes. Il est basé sur des réactions simples comme les réactions de formose qui utilisent de petites molécules qui pénètrent par diffusion passive.
- Le scénario arrive jusqu'à la synthèse des acides gras. Ce qui permet alors la reproduction par fission des liposomes. Quand l'évolution moléculaire aboutira dans certains liposomes à la réplication de l'ADN, celle-ci pourra alors se synchroniser avec la reproduction de ces liposomes.
- Synergie entre cohésion mécanique et L-aas: Le scénario que je décris ici met en oeuvre la synergie entre les L-aas et la cohésion mécanique. Deux cercles vertueux s'établissent alors:
- La tête hydrophile comporte 2 bras, le fixe celui qui contient le glycérol estérifié à 2 acides gras, et le mobile qui porte un cation (sérine, éthanolamine, choline) ou un glycérol qui sont attirés par l'anion phosphate d'un PLD voisin. Dans cet article je démontre que la chiralité du bras fixe est déterminée par l'encombrement stérique des liaisons esters (L-glycérol-3P) pour les bactéries et celui des liaisons éthers (D-glycérol-3P) pour les archées.
3.5.14 Paris
Le concept global de l'origine de lavie.
Dans l'esprit historique que j'ai adopté jusqu'ici.
- Poser les pbs restants concernant l'article chiralité.
- un pseudo-article ( pour poster ): poser les pbs de la chimio-osmose (juste à côté des autres comme chiralité)
- La pression hydrostatique ---> contrainte....
- Liposome: auto-organisation par une force (vdW+H) impose une contrainte pour l'extérieur ----> organisateur + délégation protéines (force squelette). Les éléments sont des organisateurs ( et ont été organisés par astro). Auto-organisation ADN par une force H latérale + aromaticité + vdW, n'impose pas de contrainte directement ----> délégation ARN.
- Liposome: action/réaction -----> quantique
glycolaldéhyde = ga ; acétaldéhyde=aca. ga ---> DHA <-----> DGA + DHA -------> R ; DGA + ga -------> N ; DGA + aca ------> dN ; DHA ==> LGA (mb.).
B + DGA + ga -----> N -----> P2N ;
B + DGA + aca ====> dN ---> P1dN qui aide les réactions N -----> P2N; Les P2N font évoluer la voie de néo-synthèse du ribose R qui ajouté à une base B donne N puis P2N qui active la voie de néo-synthèse du ribose R. ( Les bases sont sensées être dans la membrane par hydrophobicité et présentent un NH fonctionnel qui réagirait avec DGA−P ou DHA−P (DHA qui adopterait la forme DGA par tautomérie énol-cetone) comme LGA−P ou DHA−P s'hydrogénisent et s'estérifient en même temps aux 2 acides gras de la membrane. A cet ensemble se lie ga ou aca pour donner respectivement N (B-sucre-P) et dN d'où la notation P2N pour écrire B-sucre-PPP)
Voie de néo-synthèse du ribose R: DGA + DHA - - - - - - > R + B -------> N -----> P2N. (voir parenthèse ci-dessus).
4.5.14 Paris
2ème lecture de continuité:
- comment passer de EM, basée sur le liposome, à ED basée sur l'ADN.
- quelles sont les contraintes physiques qui permettent de passer d'une organisation à 2 dimensions sans info et dont les molécules sont libres (liposome) à une organisation semi-rigide à une dimension (ADN) contenant une info (où les molécules sont fixées)?
- Liposome: 2 forces principales, vdW + ionique et éventuellement H (du glycérol du PLDg); , les ions en zwitterions qui se neutralisent.
- L'ADN: toujours les 3 forces , toutes aussi importantes les unes que les autres + un champs d'électrons pi en résonance.
- la force ionique ----> poly-anions neutralisés par les mono-anions alcalins
- les liaisons H font la fermeture
- et les forces de vdW chassent l'eau.
- Les forces agissantes :
- entre liposome et protéines ce sont les zwitterions qui agissent;
- dans les protéines ce sont les liaisons H du squelette qui agissent; Les radicaux n'agissent pas par liaison hydrogène (ils n'en ont pas ou très peu) mais par leurs ions, leur chaine aliphatique et par les électrons pi pour contraindre la forme de la protéine.
- dans l'ADN ce sont les liaisons H des bases qui agissent. Les bases sont les radicaux des mono-mères alors que le squelette porte les ions.
- Chélation de nucléotides par les peptides.
- La quantique du liposome.
- ARN/Protéine ----> relation catalytique, labilité de l'ARN.
- ADN/Protéine -----> relation spéciale ----> transcription.
- C/L:
- Thermo= définitions, surfaces minérales (littérature)
- Liposome: plan/sphère limitée. Surface minérale avec PO4 mobile, zwitterion non ionique/SiO4. ----> double C/L. (vésicule?).
- ADN: cristal, voir les 3 forces et résonance pi ci-dessus.
- Organisation:
5.5.14 Paris
Chiralité:
- dipôle ok avec chiralité;
- tête horizontale ok avec chiralité, mais fréquence élevée des mouvements de la tête , sans donner le sens.
- Une idée puisqu'on travaille à haute pression: est-ce que la pression hydrostatique peut intervenir pour réduire cette fréquence? Est-ce que la p.h peut contraindre la chiralité L dans la vésicule pour l'agrandir? Est-ce que la chiralité L peut contrer la p.h externe et rendre cette pression plus faible à l'intérieur? A-t-on mesuré la p.h à l'intérieur du liposome?
- Une idée pour étudier l'action à distance à travers le liposome de la chimio-osmose prébiotique: La p.h rendrait plus compact et plus solide le liposome, augmentant ainsi ses propriétés quantiques pour son interaction avec l'extérieur.
- Ces 2 idées m'ont poussé à faire des recherches sur les hautes pressions et les propriétés des milieux condensés où l'action à distance du champs électrique n'a pas de sens, puisqu'il n'y a pas de vide. Ce qui m'a amené à étudier aussi l'hydratation des ions dans les solutions aqueuses, à reconsidérer le pb de la ségrégation entre Na+/K+ et la notion de contrainte physique qui remplacerait la notion de force de la physique classique. En effet dans un milieu condensé il n'y a pas de vide, mais aussi il n'y a pas de point de contact. Les nuages électroniques répondent à la physique quantique et par là n'ont pas de frontière bien délimitée: une molécule donnée dans un milieu condensé est contrainte par les molécules voisines qui l'entourent. Cette contrainte est équilibrée par la répulsion entre les nuages électroniques.
- C'est en généralisant cette notion de contrainte à toute structure et aux interfaces entre ces structures que j'ai développé un concept global pour l'évolution moléculaire.
6.5.14 Paris
Un concept global pour l'évolution moléculaire
Principe de C/L; auto-organisation; directionnalité/réversibilité; A/R (interactions entre macro-molécules) ; continuité; reproduction.
Organisation = organisation, organisateur, auto-organisation.
Continuité = 1 processus dans l'actuel, complexe, débute simplement à un certain moment dans l'évolution moléculaire et continue à se produire et à se complexifier dans cette évolution.
- Cas des facteurs de transcription: ils sont produits par la membrane par réaction au milieu extérieur puis ils agissent sur l'ADN ( sans catalyse, pour permettre la transcription ). Cette situation peut se produire à l'initialisation du métabolisme avec les pseudo-protéines ( aas piégés ou séquestrés, voir chimio-osmose et chiralité) qui encapsulent les dN par liaisons H, puis double brins, ouverture et appariement avec les N, ligases ----> ARN.
- Cas de la chimio-osmose et la formation des canaux, toujours par réaction au milieu extérieur.
- Cas des ARN: l'ARN est toujours encapsulé par des protéines, mais ces protéines ne sont pas la conséquence d'une action externe, elles sont dans le cytoplasme. Cet ensemble participe au métabolisme central. A l'initialisation du métabolisme ce sont les processus qui se développent sur la paroi interne qui produisent ces protéines, comme je l'ai supposé avec la séquestration des aas ( voir chiralité ). Mais dans ce cas les mono-ARN ne sont pas produits seulement par le métabolisme, ils sont en plus réunis par appariement sur l'ADN et peuvent produire des oligomères par des pseudo-ligases.
Si dans le cas de l'ADN la complexification est unidirectionnelle, dans le cas des ARN elle peut prendre au moins 4 directions principales:
- 1 ou 2 nucléotides encapsulés par des protéines joueront le rôle de coenzymes: ATP, GTP, NAD;
- les équivalents des tRNAs ( 70 pb environ) évolueront en tRNA et synthase;
- l'équivalent de RNAr évoluera en ribosome;
- l'ensemble + 1 RNA quelconque évoluera en traduction.
- Cas du métabolisme
- Ce n'est pas le cas de la reproduction des liposomes, ils n'évoluent pas vers la réplication.
- Les toutes 1ères synthèses des mono-nucléotides. C'est la continuité entre EM et ED.
- métabolisme actuel: DGA−P + aca <−−−> dR−1P, 4124 5427 ! dR−1P + B <−−−> dB−ose−P, 2421 3135 ! dB−ose−P + PP <−−−> dBTP, 2476 2748 ! DGA−P <−−−> Glycérate−3P, 121.12 Eco 273 Eco ! Glycérate−3P ==> Labo. !!! DGA−P <−−−> DHA−P, 5311 ! DGA−P + DHA−P -----> R−5P -----> PRPP, B1 Zn ! PRPP -----> Bases, Hist ! R−5P -----> B−ose−P ! B−ose−P + PP -----> BTP.
Est-ce que le ga est utilisé en prébiotique? où l'on a DHA−P H2 −−−> PLD ! LGA−P + H2 −−−> PLD ! mais est-ce qu'on a DGA−P + ga −−−> R−1P ??
Dans le métabolisme actuel le ga est utilisé dans: DHA−P + ga <−−−> L−xylulose−1P, 412.19 Eco ! L−xylulose−1P <−−−> D−Ribulose−5P ! pyruvate + ga <−−−> ---, 412.18 412.28 ! folate + ga <−−−> ---, 412.25 Eco ! 121..21 Eco, 111.77 Eco, 1438 EtN. - Reacion de formose: fa + fa <−−−> ga ! ga + fa <−−−> GA (L,D) <−−−> DHA ! DHA + ga <−−−>Ribulose <−−−> Ribose.
- Prébiotique:
- Formose + phosphorylation
- Actuel sans enzymes
- Alternative: DGA−P + B −−−> B−DGA−P sur membrane ! B−DGA−P ( sur membrane ) + aca −−−> dB−ose−P ! B−DGA−P ( sur membrane ) + ga −−−> B−ose−P ! dB−ose−P (ou B−ose−P) + PP −−−> dBTP (ou BTP) ! (5 réactions) DGA−P + DHA−P ----> B, Hist; C'est la voie évolutive. Dans alternative il y a directionnalité par la membrane et déplacement thermodynamique: dBTP avant BTP. BTP fait évoluer la voie évolutive et d'où le métabolisme.
- métabolisme actuel: DGA−P + aca <−−−> dR−1P, 4124 5427 ! dR−1P + B <−−−> dB−ose−P, 2421 3135 ! dB−ose−P + PP <−−−> dBTP, 2476 2748 ! DGA−P <−−−> Glycérate−3P, 121.12 Eco 273 Eco ! Glycérate−3P ==> Labo. !!! DGA−P <−−−> DHA−P, 5311 ! DGA−P + DHA−P -----> R−5P -----> PRPP, B1 Zn ! PRPP -----> Bases, Hist ! R−5P -----> B−ose−P ! B−ose−P + PP -----> BTP.
Le principe C/L : Cette notion de contrainte n'est pas la propriété de la pression hydrostatique seulement. On la retrouve pour une même pression à l'interface entre un solide et un liquide. Les molécules du liquide, en contact avec la surface solide, sont adsorbées, c.a.d fixées et donc adoptent plus ou moins l'organisation du solide. Elles sont contraintes et elles-mêmes contraignent leurs voisines qui sont plus libres qu'elles. Le phénomène d'adsorption est à la base de la catalyse par les surfaces minérales. Apparaît alors une notion plus constructive qui est celle de contrainte/liberté. Ces 2 propriétés intimement liées sont la propriété principale du vivant: Si on avait une contrainte absolue, comme on l'a pour les atomes figés d'un cristal, on aurait un minéral comme on le comprend en général. Si on avait une liberté totale, comme pour les molécules d'un liquide, on aurait un système ouvert qui n'assure aucune organisation. Le mélange de ces 2 propriétés se retrouve dans un rapport variable dans toutes les structures biologiques et notamment les liposomes. Dans le liposome les molécules qui le constituent, les PLDs, sont complètement libres en 2 dimensions, la sphère du liposome, mais elles sont contraintes dans cette sphère double minérale. Dans l'ADN, l'ARN et les protéines les monomères sont fixes dans une dimension mais peuvent se mouvoir plus ou moins autour de cette dimension. La structure biologique qui se rapproche le plus du cristal (du minéral) c'est l'ADN sous sa forme de double brin: l'appariement des 2 brins se fait par liaison H nettement plus forte que les liaisons vdW et ioniques, mais peuvent se défaire et se faire sans réorganiser la structure. Grâce aux liaisons hydrogènes on peut cristalliser, en labo, les protéines et les acides nucléiques. Les virus sont des quasi cristaux de protéines et d'acides nucléiques mais se font et se défont grâce aux liaisons H aussi.
Le principe d'organisation (auto?):
- auto-évolution
- organisation imparfaite, cristal (ion +liaison covalente), liposome (vdW), protéine (liaisons H sur squelette), ADN (liaisons H sur radicaux), ARN (peu de liaisons H)
- Évolution par délégation: liposome aux protéines, ADN à l'ARN
- Organisation + C/L donne: un liposome à 2 dimensions, un ADN à 2 types de paires de bases, des protéines(dipôles) et des ARN (apparié par morceaux) d'une grande variété.
- L'organisation crée de nouvelles forces donc de nouvelles contraintes pour son environnement: ADN (résonance), liposome (zwitterions, osmose), protéines (dipôles, feuillets beta), ARN (structuration de la catalyse), interactions entre macro-molécules.
Le principe de C/L s'applique aux éléments d'une structure donnée et celle-ci par sa contrainte sur les molécules avoisinantes synthétise d'autres molécules qui pourraient former une autre structure avec une nouvelle contrainte différente de la précédente.
- **
- Les molécules synthétisées, dans la plupart des cas, ne sont pas organisées en une nouvelle structure. Dans le cas d'un système fermé comme le liposome les molécules libres synthétisées par les structures organisées sont canalisées par les contraintes de ces structures et constituent le métabolisme central. Par leurs contraintes les structures organisées font partie donc du métabolisme central.
- Le liposome dont les acides gras sont synthétisés par les contraintes des surfaces minérales (postulat de la poche de pétrole prébiotique) est la seule structure auto-organisée par les petites molécules. Il en contient quelques millions. Les forces d'organisation sont les plus faibles, les forces de vdW. Comme on l'a vu dans la chiralité prébiotique ce sont les contraintes des têtes carboxyliques du liposome ( ou de la vésicule) qui initialisent le métabolisme et modifie ces têtes en zwitterions. Ces surfaces partiellement ou totalement zwitterioniques piègent ou séquestrent les aas de la soupe prébiotique. Ces aas pénètrent dans la membrane et constituent les 1ères protéines membranaires dont les canaux, peut-être sous forme de groupements d'aas non liés mais contraints par et dans la membrane.
- La chimio-osmose est le moteur de l'évolution moléculaire.
8.5.14 Vendôme
Le principe d'organisateur:
** Cette faculté d'une structure à organiser son environnement je l'appelle le principe d'organisateur. Ce principe se manifeste avec toute structure moléculaire mais aussi avec les atomes isolés comme les ions mono-atomiques et les petites molécules de quelques atomes comme le P (phosphate). Le pouvoir organisateur dépend de la complexité et de la taille de la structure organisatrice. Il augmente par exemple avec le nombre atomique dans une colonne du tableau de Mendelev. K+ par exemple devrait être plus organisateur que Na+ sur H2O parce qu'il a 8 électrons en plus.
23.5.14 Paris
Les 5 zones de la chimio-osmose
- Évolution de la face externe du liposome:
La surface du liposome, faite de 10 millions de têtes zwitterioniques environ, favoriserait la pénétration des acides aminés et des acides alpha-hydroxylés dans la membrane et leur regroupement sur cette surface.
- Cohésion du liposome: Cependant l'encombrement stérique des têtes hydrophiles, imposé par le rapprochement des chaînes aliphatiques, oppose une forte contrainte à la pénétration. Et cet encombrement est renforcé par la longueur du bras mobile, l'éthanolamine par exemple, qui arrime un phospholipide à son voisin. C'est ce qui fait d'ailleurs la solidité et la cohérence du liposome. Une méthylamine à la place de l'éthanolamine ne permettrait pas l'arrimage et une propylamine le rendrait trop lâche.
- Configuration des acides aminés: La cohésion du liposome devrait imposer une configuration stérique aux acides aminés et aux acides alpha-hydroxylés qui pénètrent dans la membrane. Cette configuration concerne leur longueur et leur chiralité.
- Synergie structure/chimio-osmose: Nous voyons ainsi qu'à la face externe et même qu'à la face interne le liposome intervient par sa structure même, sans tenir compte du processus chimio-osmotique auquel elle s'ajoute. Le processus chimio-osmotique prébiotique, même si son action peut paraître infiniment plus faible que celle du processus biotique, il évolue néanmoins dans un milieu réactionnel structuré par et avec le liposome et se renforcent mutuellement.
- Évolution de la zone hydrophobe de la membrane:
Elle va hériter des molécules et de la réactivité chimique du milieu extérieur qui, dans la poche prébiotique, la soupe prébiotique.
- Intégration des acides aminés: L'intégration des petites molécules polaires ou même ionisées par les têtes hydrophiles dans la membrane va les obliger à prendre des configurations spatiales propres aux milieux aliphatiques. Les groupements d'acides aminés et des acides alpha-hydroxylés, accompagnés éventuellement de catalyseurs minéraux chélatés, peuvent devenir des centres catalytiques ou des canaux ioniques.
- Les molécules hydrophobes: Ces molécules, comme les hèmes, les hydroquinones et les bases nucléiques s’intercalent facilement dans la membrane et peuvent présenter de part et d'autre des fonctions chimiques qui seront modifiées chimiquement par les milieux intérieur et extérieur, ou bien serviront comme coenzyme aux groupements d'acides aminés intégrés dans la membrane.
- Transduction du potentiel électrique: Les molécules hydrophobes qui s'étendent sur toute l'épaisseur de la membrane peuvent servir de transducteur du potentiel électrique.
- Évolution de la face interne du liposome:
- Elle la même structure zwitterionique que la face externe mais une partie des têtes hydrophiles peuvent être des sérines responsable en partie du potentiel électrique entre les 2 faces.
- Elle va hériter des molécules triées par les 2 zones précédentes. Les acides aminés seront peut-être plus homogènes, surtout en ce qui concerne la chiralité.
- Elle va faire pénétrer dans la membrane les molécules propres au milieu intérieur, notamment les petits acides aminés qu'on a vu dans la partie du poster consacrée à l'initialisation du métabolisme dans la poche de pétrole prébiotique et qui sont Gly Ala Ser Asp Glu ainsi que les acides aminés dérivés de ces derniers par le métabolisme naissant.
- En contact avec le milieu intérieur, elle va subir sa réactivité chimique et son organisation.
- Le milieu intérieur:
- Un milieu fermé: Il est fermé et petit, limitant la diffusion et favorisant l'organisation.
- Des réactions chimiques limitées pendant l'initialisation du métabolisme: Dans l'hypothèse de la poche du pétrole prébiotique, la soupe prébiotique contient de petites molécules qui diffusent facilement à travers la membrane. Ce sont H2, le formaldéhyde, l'acide formique, l'acétaldéhyde, NH3 et CO2 et sont à l'origine de la réaction de formose, qui produits les trioses, et des réactions qui produisent les petits acides aminés ( voir ci-dessus ).
- Le potentiel électrique devrait modifier l'ionisation des acides et des bases faibles.
- La séquestration du phosphate: A l'intérieur le phosphate sera intégré dans le métabolisme naissant et plus tard dans les acides nucléiques. C'est ce que j'appelle la séquestration du phosphate. Le phosphate est poly-atomique, gros, ionisé aux pHs biologiques et peut porter 1 à 3 charges négatives. Il devrait entrer beaucoup plus difficilement que les ions mono-atomiques. Or sa vitesse de diffusion à travers la membrane est équivalente à celle de K+ et Na+. L'hydrolyse des têtes hydrophiles, déséquilibrant les phospholipides, rend possible le passage de ces phospholipides d'un feuillet à l'autre, par le processus connu appelé flip-flop. Ce qui expliquerait sa vitesse de diffusion.
votre commentaire
-
Par mekali le 14 Mars 2014 à 12:02
18.12.13 détricotage préparation en cours 18.12.13 panachage R/dR idée ok 8.1.14 gradient du vivant idée sur papier à faire. ok 13.1.14 gradient du vivant idée sur papier à faire. ok 15.1.14 les forces fondamentales à coté ok 23.1.14 2ème détricotage préparation suite ; sur papier à faire ok 28.1.14 codon-anticodon idée ok 29.1.14 2ème détricotage suite préparation suite ; sur papier à faire ok 30.1.14 zéolites pour prebiotic petroleum idée ok 31.1.14 prebiotic petroleum, réecriture papier wiki, à faire ici 4.2.14 prebiotic petroleum, réecriture papier wiki, à faire ici 6.2.14 groupes des aas papier à faire ok 11.2.14 abstract pour prebiotic petroleum papier wiki, à faire ici votre commentaire
-
Par mekali le 13 Septembre 2012 à 21:51
Initialisation du métabolisme dans le pétrole abiotique
début de rédaction dans wikiversité 150611
Sommaire
- 1 Introduction
- 2 État de la recherche sur la poche de pétrole abiotique
- 3 Hypothèses de travail pour une poche de pétrole abiotique
- 4 Initialisation du métabolisme dans les poches de pétrole abiotique analogues à celles du pétrole fossile
- 5 Références
Introduction
Ce travail se veut une recherche théorique sur l'évolution moléculaire qui se produit dans le pétrole abiotique et qui pourrait aboutir à l'émergence de la vie. Une poche de pétrole abiotique analogue à celle du pétrole fossile constitue un environnement idéal pour l'étude de l'origine de la vie. C'est un milieu fermé, stable, riche en molécules organiques et en catalyseurs minéraux, évoluant et remontant à un rythme géologique des profondeurs de la croûte terrestre à la surface. En passant progressivement des conditions physico-chimiques extrêmes des profondeurs à celles de la surface, compatibles avec la vie, toute évolution moléculaire qu'on puisse imaginer y est possible.
Le pétrole synthétique, pourrait être utilisé dans une expérimentation abiotique. Mais il est synthétisé par le procédé industriel de Fischer-Tropsch à partir du gaz de synthèse, CO+H2, lui-même synthétisé soit du coke (C+H2O) soit du gaz naturel(CH4+H2O) tous deux d'origine fossile. Pour arriver à un pétrole abiotique, analogue au pétrole fossile, il faut que sa synthèse se fasse en présence des éléments N P S, avec des molécules d'origine abiotique, en profondeur dans la croûte et qu'il remonte imbibé dans une roche poreuse exempte de toute molécule d'origine biogénique.
Avant d'étudier l'initialisation du métabolisme dans ce contexte, nous allons établir l'état des connaissances sur les processus géochimiques produisant du pétrole abiotique accompagné des éléments N P S sous forme abiotique aussi.État de la recherche sur la poche de pétrole abiotique
Ce sera plutôt l'état de la recherche sur les éléments de la poche qui la composeraient. Car aucun travail n'a été fait jusqu'à maintenant à son sujet. Ces éléments sont les hydrocarbures et l'eau (H C O), le phosphate libre (P) et l'ion ammonium(N). Nous n'étudierons pas le soufre (S) spécialement, car ses espèces moléculaires qui interviennent dans le vivant, sulfures, sulfates et l'élément S lui-même sont omniprésents dans la lithosphère et l'hydrosphère.
Pour l'azote, NH4+ est l'espèce la plus présente en profondeur, aux hautes pressions et aux hautes températures, car dans les minéraux il remplace l'ion potassium K+. Au contraire, en surface et aux pressions et températures de la poche de pétrole fossile, ce sont les nitrates qui représentent l'azote, car l'ammonium est utilisé directement par les bactéries. Le diazote N2 dissout dans les eaux interstitielles à 11 ppm, peut être réduit en NH4+ en présence de H2 dans les fluides hydrothermaux ce qui n'est pas le cas en surface et dans la poche de pétrole fossile.Sur le terrain
Le phosphate
- Le phosphore est présent à 700 ppm molaire( 1050 ppm massique [1]) dans la croûte terrestre sous forme, la plupart du temps, d'ions phosphates intégrés dans les apatites insolubles et à 0,04 ppm molaire (2µmol/kg, Broecker & Peng [2] 1982) dans l'eau des océans sous forme de phosphates libres.
- Aux températures généralement atteintes dans les magmas, 1000-1400 °C, une fraction des ions phosphates dans le liquide magmatique se polymérise pour former des polyphosphates. Yamagata[3] et al. (1991) a démontré que des polyphosphates sont présents dans les solutions hydrothermales à une concentration de l'ordre du micromolaire.
- D'après Arrhenius[4] et al. (1997), dans l'eau de mer (Mg/Ca ~ 5 et pH < 8.5 ) les phosphates provenant du lessivage des terres émergées, précipitent sous forme d'un solide amorphe d'hydrogénophosphates de Ca et de Mg (brushite[5], CaHPO4.2H2O et newberyite[6], MgHPO4.3H2O).
Ce solide cristallise en whitlockite[7], HMgCa9(PO4)7, capable de donner des oligo-phosphates dans les conditions de température et de pression du métamorphisme de contact ou de celles des sources hydrothermales de surface. D'après Arrhenius [4] , aujourd'hui, les êtres vivants utilisent les hydrogénophosphates et les transforment en apatites. Et aux origines de la vie, les oligophosphates seraient très disponibles.
- On trouve la whitlockite comme minéral secondaire dans les pegmatites granitiques et en abondance dans les environnements extraterrestres : les météorites, sur la Lune et sur Mars.L'ammonium
- L'azote est présent à 35 ppm massique(50 ppm molaire) dans la croûte terrestre sous forme d'ions ammonium remplaçant les ions potassium dans les roches ignées (Hall [8] 1999, Stevenson [9] et al. 1962) .
D'après Hall la majorité de cet ammonium est d'origine sédimentaire, mais une partie est d'origine magmatique. L'origine sédimentaire est incorporée majoritairement par altération hydrothermale, le reste par métamorphisme de contact. L'ammonium des roches ignées peut être extrait par pyrolyse sous vide à 1000 °C sous la pression standard (Hall [10] et al. 1993). Ce qui correspond peut-être aux conditions du manteau supérieur.
- Les nitrates, très rares, sont représentés par les évaporites en surface. Les nitrates dissouts ne représentent que 0,6 ppm molaire (30 µmol/L, Broecker & Peng [2] 1982) dans les océans.
- Dans l'eau de mer et les eaux interstitielles le diazote est la forme prédominante avec 11 ppm molaire (590 µmol/L, Broecker & Peng [2] 1982 ); le diazote peut être réduit en NH3 par H2 des fluides hydrothermaux.Le pétrole abiotique
C'est essentiellement le processus de serpentisation qui a été étudié sur le terrain , pour la recherche du pétrole abiotique. Et notamment au niveau des dorsales océaniques et aux premiers kilomètres de profondeur des zones de subduction.
- Dorsales océaniques.
- Les dorsales océaniques s'étalent sur 60 000 km et sont parsemées régulièrement de sources hydrothermales qui produisent sur de longues durées H2S, CH4, H2, N2 et CO2. Ces 5 gaz sont refroidis brusquement à 2 °C, la température des fonds marins, à la sortie des sources et se retrouvent sur le plancher océanique, sous des pressions de 100 à 400 bars et plus, où ils peuvent former des clathrates de gaz. En s'éloignant de l'axe des dorsales, ils seront recouverts petit à petit de sédiments contenant les hydrogénates de phosphate qu'on a vu ci-dessus.
- Voici en condensé, les résultats des campagnes de mesures faites dans 7 sources hydrothermales de l'Atlantique, d'après Charlou [11] et al. (2002), table 1.
- Tableau 1. Les gaz produits sur les dorsales atlantiques.
-
Gaz
mmol/kgnombre
de sitesintervalle Eau de mer CO2 6 5.2 - 28 2.30 N2 6 .9 - 3 .59 H2S 7 .5 - 11 0 CH4 7 .023 - 2.63 .0003 H2 7 .020 - 16 .0004 δ13C CH4
en ‰6 -8 . -19,6 -
- Au site Rainbow (Charlou [11] et al. 2002, table 3) les mesures ont été faites en 1997, 1998, 1999 et 2001. Elles ne varient guère .
- Par ailleurs la mesure du fractionnement isotopique du 13C pour CH4 (δ13C) laisse penser à une origine abiotique. Le méthane serait synthétisé par le processus Fischer-Tropsch .
- L'origine abiotique du méthane a été étudiée plus en détail par Proskurowski [12] et al. (2008), en montrant l'augmentation du δ13C pour C2H6 et C3H8 avec le nombre de carbones, conforme à la théorie du fractionnement abiotique.
- Zones de subduction
- Au contact des 2 plaques en subduction, les processus géochimiques sont multiples et complexes, dus aux forces gigantesques mises en jeu, sans commune mesure avec ce qui se passe sur les dorsales océaniques. Au niveau des fosses océaniques les pressions peuvent atteindre le kbar et la température n'est que de 2 °C.
- Aux premiers km de profondeur ce sont donc des processus à basse température et haute pression qui sont en action. Le faciès schiste-bleu se situe ainsi à 15-18 km de profondeur avec des pressions supérieures à 6 kbar et des températures entre 200 et 500 °C. Les roches ultramafiques vont se métamorphoser en lizardites, qui sont des serpentinites. D'après la synthèse sur les serpentinites faite par B.W. Evans [13] (2004), la serpentinisation peut se produire entre 50 et 300 °C de température et dans une large gamme de pression, de 0.1 kbar à 10 kbar (figure 5 [13]), pour donner de la chrysotile[14] ( Mg3Si2O5(OH)4 ) et de la lizardite[15] ( Mg3Si2O5(OH)4 ) avec production de H2 et de magnétite.
- Au-delà de 18 km de profondeur, la serpentinisation des roches ultramafiques donnera de l'antigorite[16] ( (Mg,Fe++)3Si2O5(OH)4 ) sans production de H2 ni de magnétite à une température entre 400 et 600 °C (Evans [17] 2010).
- Pour les zones de subduction, ce sont là les seuls résultats qu'on puissent déduire indirectement, de l'étude des roches et des anomalies magnétiques et sismiques au niveau de la zone avant de l'arc volcanique.
- Sinon de nombreux phénomènes en surface laissent penser qu'il y aurait production d'hydrocarbures, mais dont on ne sait pas, jusqu'à maintenant, s'ils sont de nature fossile ou abiotique. Ce sont :
- Les volcans de boue à l'avant de l'arc, qui émettent beaucoup de méthane.
- Les hydrates de gaz repérés par de nouvelles techniques tout le long des zones de subduction, d'une longueur équivalente aux dorsales océaniques. Ces gisements ne sont pas encore exploités.
- Les gaz de schiste et les schistes bitumeux, roches issues des zones de métamorphisme.
- Les gaz volcaniques qui comprennent CH4, H2 et NH3.
- Il ne faut pas oublier aussi la forte circulation hydrothermale vers le haut, produite par la vaporisation de l'eau et des gaz qui imbibent la croûte océanique en subduction. Ces fluides hydrothermaux transportent notamment des ions NH4+ qui remplacent les ions K+ dans les roches métamorphiques comme on l'a vu précédemment, ainsi que du phosphate libre[3].
En laboratoire
Les expérimentations abiotiques en laboratoire sont beaucoup plus nombreuses et variées que les mesures sur le terrain, et elles concernent les 3 molécules clés que sont le méthane, l'ammoniac et le phosphate. Le rapprochement de la synthèse de ces 3 molécules dans le tableau 2, est justifié du fait que, elles et les molécules qui leur donnent naissance, c'est-à-dire H2, CO2 et N2 se retrouvent toujours ensemble dans les fluides hydrothermaux. Et cette situation est tout à fait adéquate pour la formation de poche de pétrole abiotique contenant les éléments N P S.
Les expériences rapportées au tableau 2, n'ont pas été faites toutes dans le même esprit, ni dans l'objectif de la poche de pétrole abiotique. Cependant les résultats sont compatibles avec la formation d'une poche abiotique. Les principaux enseignements qu'on peut en tirer sont :- Ces processus se produisent dans une gamme de températures étroite, autour de 300 °C ;
- La pression peut varier beaucoup pour un même processus, jusqu'à 2 ordres de grandeurs, mais reste inférieure à 10 kbar pour tous les processus.
- L'état supercritique des expériences hydrothermales permettent de les mettre en parallèle avec les procédés industriels en phase gazeuse et à faible pression. On a ajouté le vaporeformage car il peut se produire en subduction, comme l'attestent les gaz volcaniques des arcs de subduction.
- Les catalyseurs utilisés, notamment le fer, son oxyde et le nickel, sont abondants dans la lithosphère.
- Il faut noter l'importance des surfaces minérales dans les procédés industriels. Il est primordial que les expériences pour l'objectif « poche abiotiques » soient faites dans des roches poreuses adéquates.
- Les expériences hydrothermales sont motivées par l'hypothèse que l'origine de la vie s'est produite (ou se produit) au niveau des dorsales océaniques. Or il est évident que la trempe immédiate de H2, CO2 et N2, dans l'eau froide du plancher ne devrait pas du tout favoriser les processus de type Fischer-Tropsch et la réduction du N2 par les processus de type Haber-Bosch. Mais les résultats sont intéressants car les expériences sont faites à haute pression et haute température qui sont les conditions des zones de subduction. De même c'est intéressant pour les processus se déroulant dans les hydrates de gaz, qu'on a vu précédemment, quand ils seront recouverts de quelques km de sédiments tout en subissant la chaleur du plancher qui se refroidit.
- McCollom [18] a pu démontrer, ligne e.j du tableau, que la discrimination entre biotique et abiotique sur la base du fractionnement des carbones 12 et 13 n'était pas valable pour le méthane. C'est pour ça que pour démontrer la nature abiotique des hydrocarbures au niveau des dorsales, Proskurowski [12] a dû le faire pour les chaines d'hydrocarbures supérieures. Cette remarque a une grande importance pour la suite, car si l'origine de la vie s'était faite dans le pétrole abiotique, alors les êtres issus de ce pétrole auraient pu transformer, ou seulement enrichir, avec leurs membranes, les hydrocarbures abiotiques en hydrocarbures biotiques. De là l'origine biotique des pétroles dits fossiles.
- Ces expériences doivent être refaites dans l'hypothèse de la poche de pétrole abiotique et notamment pour les phosphates, aux mêmes pressions que les autres expériences.
- Tableau 2. Les processus abiotiques
-
Processus réactions Produits Température °C Pression bar a Vaporeformage CH4+H2O CO H2 800-900 25 b Serpentinisation dorsales, plancher FeO+H2O H2 N2 CH4 CO2 50-300 100-10000 c Serpentinisation subduction 15km FeO+H2O H2 (N2 CH4 CO2 ?) 200-500 6000 d Fischer-Tropsch gaz CO+H2 Pétrole dont acides gras 220-350 25-45 e,j Fischer-Tropsch hydrothermal CO2+H2 Pétrole dont acides gras 250 325 f Haber-Bosch gaz N2+H2 NH3 500-600 200-300 g Haber-Bosch hydrothermal N2+H2 NH3 700 1000 h Haber-Bosch hydrothermal N2+H2 NH3 200 55 i Arrhenius chauffage MgHOPO3+H2O OH(H2PO3)nH 100-500 1
- a. Vaporeformage: [19]
- CH4 + H2O = CO + H2;
- T= 800-900 °C, P= 25 bars, catalyseur: Ni, CuO, Zn, surfaces minérales.
- Serpentinisation
- (Mg,Fe)SiO4 + H2O = Mg3Si2O5(OH)4 + Mg(OH)2 + Fe3O4 + H2 , ou bien
- olivine + eau = serpentine + brucite + magnétite + hydrogène et se résume à
- 2(FeO)roche + H2O = (Fe2O3)roche + H2
- b. Serpentinisation dorsales, plancher
- Reproduction de la serpentinisation en laboratoire et modélisation :
- McCollom [20] 2009, modélisation ;
- B.W. Evans [17] 2010, expérimentation.
- Température de 50 à 300 °C, Pression de 100 bars à plusieurs kbar.
- Holm [21] 2006 : considère que sur le plancher la serpentinisation se fait de façon continue à des températures autour de 150 °C.
- c. Serpentinisation subduction à 15km
- Evans [13] (2004), (wikipedia anglais) .
- Température de 50 à 300 °C, Pression de 6 kbar.
- Processus Fischer-Tropsch
- H2 + CO = CH4 + CnH2n+2 + CH2O + CH3OH + C2H4 + C2H5OH + (alcools et acides gras à longues chaines).
- d. Procédé industriel phase gaz : Kreutz [22]et al. 2008
- T= 220-350 °C, P= 25-45 bars; catalyseur: Fe, surfaces minérales.
- e. Conditions hydrothermales, McCollom [23]1999:
- T= 175 °C, P= 325 bars; catalyseur: montmorillonite + alumine.
- j. Fractionnement isotopique, McCollom [18]2006:
- Démonstration du fractionnement non discriminant des isotopes 12 et 13 du carbone, processus Fischer-Tropsch 23 en conditions hydrothermales, T= 250 °C, P= 325 bars; catalyseur: Fe.
- Processus Haber-Bosch.
- N2 + 3 H2 = 2 NH3
- f. Procédé industriel phase gaz Haber-Bosch:
- T= 500-600 °C, P= 200-300 bars, catalyseur Fe, surfaces minérales.
- g. Conditions hydrothermales, Brandes [24]1998:
- T= 300-800 °C, P= 1000 bars, catalyseur Fe : pour un rapport H2O/Fe = .5, production de 17% de NH3 à 700 °C
- T= 500 °C, P= 1000 bars, catalyseur Fe3O4 dans HCO2H : production de 0.6% de NH3.
- h. Conditions hydrothermales, Smirnov [25]2008:
- T= 200 °C, P= 55 bars, catalyseur Fe.
- i. Production des polyphosphates et leur concentration par les surfaces minérales.
- Arrhenius [4] 1997:
- Newbeyrite, Mg(OHPO3).3H2O donne des oligo-phosphate jusqu'à 9 phosphates. Pression 1 bar, chauffage de 100 à 550 °C
- de même avec brushite, Ca(OHPO3).2H2O et withlockite, HMgCa9(PO4)7.
Hypothèses de travail pour une poche de pétrole abiotique
Étant donné la profusion de H2 produit par la lithosphère : dorsales océaniques, zones de subduction et même dans les eaux interstitielles des roches, on est obligé de tenir compte, dans la quête de l'origine de la vie, du méthane et du pétrole abiotiques qui pourraient être produits à partir de H2 et du CO2 de la serpentinisation.
On peut considérer ainsi 3 types de méthane dans la lithosphère suivant leur origine :- Le méthane issu de la serpentinisation à des pressions inférieures à 10 kbar et une température inférieure à 300 °C, en profondeur. Les clathrates de gaz repérés dans les zones de subduction contiendraient ce méthane.
- Le méthane biogénique de la surface, produit par les procaryotes méthanogènes à partir de la biomasse : feu follet, clathrates des marais, lacs, permafrost et plateaux continentaux des océans.
- Le méthane abiotique produit au niveau du manteau supérieur à partir de carbonates et d'eau à des pressions supérieures à 50 kbar et à une température autour de 1500 °C. Ce méthane a été produit en laboratoire seulement (H.P Scott [26] et al., J.F Kenney [27] et al.). On n'a pas pu, jusqu'à maintenant, montrer son existence sur le terrain. Il est supposé se transformer en partie en pétrole et remonter par les failles ou le long des plaques de subduction.
Je ne traites pas ici le méthane des grandes profondeurs (le type 3), car il concerne la polémique du pétrole abiotique dans le contexte de la production industrielle.
Pétrole issu de la serpentinisation
Qu'en est-il du pétrole produit par le processus FTT en même temps que la serpentinisation? Au niveau des dorsales, il n'est produit qu'à l'état de traces alors que le rapport H2/CH4 peut-être très élevé. Que devient cet hydrogène s'il n'est pas utilisé pour la synthèse du pétrole?
-
- Sur les flancs des dorsales océaniques: diagénèse des clathrates de gaz.
Une hypothèse émise par les spécialistes des dorsales, dont J.L Charlou [28] (Ifremer: Serpentinisation et synthèse inorganique d'hydrogène, méthane et hydrocarbures le long de la dorsale médio Atlantique), serait la formation d'hydrates de gaz de méthane piégés dans les sédiments sur les flancs des dorsales. Ce méthane serait produit directement par serpentinisation grâce à une convection hydrothermale à travers la croûte fracturée. Développons un peu cette hypothèse.
D'abord la serpentinisation ne produit pas de méthane, mais produit H2 et produit ou concentre CO2 et N2. La production de méthane se ferait par le processus FTT après. La serpentinisation se déroule entre 50 et 300 °C et à des pressions allant jusqu'à 10 kbar. Les gaz produits par les flancs s'ajoutent à ceux déjà produits par la dorsale. Sur les flancs, loin de l'axe, les fractures sont moins profondes et la température est plus basse, et la pression augmente en descendant le flanc. Cette situation permet la production de H2 à une température modérée et favorise la formation des clathrates. La production de méthane par FTT risque de diminuer plus rapidement que la serpentinisation, car elle doit se faire à une température plus élevée (voir McCollom [23] ). Cependant plus on s'éloignera de l'axe, plus la couche de sédiments s'épaissira et la pression augmentera entrainant une augmentation de la température par gradient géothermique.
Prenons l'exemple du gisement Tupi [29] découvert au large du Brésil, dans une zone sans subduction, sous 2 km d'eau et 5 km de sédiments. Avec un gradient géothermique de 30 °C/km et celui de la pression de 250 bar/km dans les sédiments, nous atteignons une température de 150 °C à laquelle il faut ajouter la chaleur apportée par la croûte, et une pression de l'ordre de 1,5 kbar. Pendant des millions d'années ces clathrates ont été soumis à des températures nettement inférieures à celle de l'optimum du processus FTT de 250 °C, et à des pression qui vont très vite dépasser les 350 bars de l'optimum pour atteindre 1 à 2 kbar. Par rapport à l'optimum, le fait que la synthèse des hydrocarbures soit exothermique, ( l'équilibre va être déplacé vers le méthane avec la baisse de la température et l'augmentation de la pression (4H2+CO2=CH4+2H2O dh=-206kj/mol ); la vitesse ralentit avec la baisse de température ; état supercritique pour les gaz et pas pour l'eau et les chaines longues), auxquelles il faut ajouter l'effet de surface du mélange sédiments/clathrates et des catalyseurs.
L'intérêt de l'hypothèse de J.L Charlou [28] , ce n'est pas tant la production de CH4, que celle de H2 et la formation des clathrates. Les études [30] [31] [32] des clathrates sont très développées actuellement pour le stockage de l'hydrogène utilisé comme source d'énergie, la séquestration du CO2 au fond des océans et la libération du méthane par réchauffement climatique. Il n'est pas question de développer ce point ici, mais ces études montrent que les conditions de température et de pression au niveau du plancher océanique, sont compatibles avec la stabilité de ces mélanges faits de sédiments et de clathrates des gaz comme ceux issus de la serpentinisation .
Il ne faut pas oublier la réduction de N2 en NH4+ par H2 par le processus Haber-Bosch dans un état supercritique (voir cours de Nancy pour la variation avec la pression, paragraphe 3 [33], et catalyseur au tantale: P. Avenier et al.2007 [34] ) .
Mais aussi la libération du phosphate des hydrogénophosphates par la chaleur et sa concentration par les surfaces minérales (Arrhenius [4] ). Ces molécules sont importantes pour notre objectif de poche de pétrole abiotique. (à développer pour distinguer des fluides hydrothermaux)
Cette hypothèse pourrait expliquer, avec la tectonique des plaques, les grands gisements de gaz et de pétrole du côté des marges passives à très grande profondeur(Tupi), les clathrates de gaz ayant migré vers les zones de subduction dans le cône d'accrétion, ainsi que les gaz de schiste par métamorphisme de ces clathrates mélangés aux sédiments, dans la zone de contact des 2 plaques.- Au niveau des zones de subductions : transformation du méthane en pétrole au contact des fluides hydrothemaux.
Au niveau des zones de subductions il a été repéré surtout des clathrates de gaz qui proviendraient, d'après moi, comme on l'a vu précédemment de la serpentinisation des dorsales. On peut toujours supposer, pour notre poche abiotique, qu'une partie de ce méthane abiotique soit transformé en pétrole au contact des fluides hydrothermaux formés à de hautes pressions et de température le long des plaques en subduction. Le processus FTT serait initialisé par vaporeformage du méthane, l'ammoniac et le phosphate sont apportés par les fluides hydrothemaux (voir chapitre ammoniac et phosphate). Les volcans des zones de subduction, terminent leur activité par un rejet d'un peu d'hydrocarbures (*). Mais la zone de la plaque subduite, entre le cône d'accrétion des sédiments et le métamorphisme de l'arc volcanique, est très réduite pour pouvoir créer de grands gisements de pétrole. Par contre ce pétrole abiotique, en petite quantité localement, mais répandu sur des dizaines de milliers de km, peut constituer autant d'opportunités pour constituer des poches favorables à l'évolution moléculaire.
- Le long des plaques transformantes:
Les hypothèses de travail pour l'expérimentation
Les principaux points à respecter pour l'expérimentation de l'initialisation du métabolisme dans le pétrole abiotique. sont donc :
- Haute pression de l'ordre de quelques kbars au début;
- Une température relativement faible, entre 50 et 300 °C ;
- Une poudre de roche qu'il faudra définir, qui simulera un dépôt sédimentaire poreux contenant des hydrogénophosphates et des catalyseurs (Fe Ni et d'autres éléments à définir suivant la littérature des procédés industriels ou expérimentaux).
- Du clathrate de gaz reconstitué contenant H2 CO2 N2 H2O
- Le H2S peut bloquer certaines réactions, aussi faut-il l'introduire petit à petit ;
- Si le produit obtenu se rapproche de la poche de pétrole fossile, on peut réduire la pression et la température en suivant l'évolution moléculaire pour repérer s'il y a initialisation du métabolisme ou non.
- Priorité à la synthèse de l'huile d'abord.
- Envisager des expériences de longues durées (ou les simuler) pour reproduire les temps géologiques et les processus de cristallisation.
Parallèle avec la théorie du pétrole fossile
31.01.2014
C'est en lisant un article sur les bactéries qui posent des problèmes dans l'exploitation pétrolière que je suis tombé sur un schéma qui représente les intéractions entre les différentes espèces de procaryotes qui se trouvent dans une poche de pétrole fossile et leur environnement (*). Ce schéma, avec la mention des gaz et des catalyseurs, ressemblerait énormément à celui issu d'une étude sur la poche de pétrole prébiotique. C'est ainsi que:
- Ces procaryotes tirent leur énergie des minéreaux et non de la photosynthèse, comme dans la poche prébiotique;
- consomment les petites molécules du pétrole et les gaz. Cependant l'ammoniac et le phosphate n'apparaissent pas à la différence de la poche prébiotique, car le phosphate est un facteur limitant et certaines espèces transforment rapidement l'ammoniac en nitrates qui est à son tour utilisé par d'autres espèces pour l'énergie et la fabrication des aas;
- des vésicules aqueuses peuvent se former dans la phase huile et des liposomes dans la phase eau et l'on est alors dans la même situation que dans la poche prébiotique pour l'évolution moléculaire prébiotique;
- Les conditions de pression, et dans une moindre mesure de température, obligent les procaryotes à s'adapter jusqu'à atteindre les conditions nécessaires à l'initialisation de l'évolution moléculaire. C'est ainsi que nous voyons qu'aux pressions extrêmes, 1.0-1.5 kbar, les procaryotes simplifient leurs protéines qui deviennent monomériques au lieu de rester multimériques, que les chaines aliphatiques s'allongent et augmentent le nombre des liaisons insaturées (*).
- Nous pouvons se trouver alors dans une situation telle qu'on ne puisse plus distinguer si le procaryote a atteint le maximum de régression pour pouvoir encore le définir comme vivant ou bien si le liposome, formé dans la poche fossile et ayant subi l'évolution moléculaire prébiotique, a atteint un stade d'évolution maximum pour qu'il passe pour un être vivant. C'est à ce niveau que se pose l'énigme des procayotes endémiques des poches de pétrole fossile: sont -ils nés du pétrole ou bien on n'a pas pu démontrer encore leur origine allochtone (*)?
La poche de pétrole fossile a l'avantage d'exister, que de nombreuses expérimentations y ont été faites pour résoudre des problèmes d'exploitation pétrolière. Ces expérimentations ont développé les techniques des hautes pressions et les conditions extrêmes de température. Cependant elles n'ont pas été faites dans l'optique de l'évolution moléculaire prébiotique. On pourrait les appliquer à la poche de pétrole prébiotique. Notamment il serait judicieux d'expérimenter une poche fossile sans procaryotes avec du phosphate, de l'ammoniac, quelques aas et des bases nucléiques pour observer l'évolution moléculaire à un stade avancé. Mais déjà au niveau de la littérature on peut étudier, toujours dans l'optique de l'évolution moléculaire prébiotique, les intéractions du milieu environnant avec le métabolisme du procaryote de la poche fossile.
Initialisation du métabolisme dans les poches de pétrole abiotique analogues à celles du pétrole fossile
On n'arrive pas à concevoir le métabolisme dans un milieu ouvert, non confiné dans un volume restreint de la taille d'une cellule. Quel que soit le réseau de réactions chimiques, qui s'établit à un moment donné dans un milieu ouvert, il est automatiquement détruit par le processus de diffusion. De même un réseau délimité par une paroi lipidique fermée, ne communiquera pas avec l'extérieur et du coup n'évoluera pas.
Beaucoup d'expériences ont été faites avec les surfaces minérales, donc des milieux ouverts, pour démontrer la catalyse et/ou la concentration de molécules organiques ou minérales comme les phosphates, les acides aminés et les acides nucléiques. Mais aucune d'entre-elles ne laisse supposer une initialisation du métabolisme, à moins d'imaginer des scénarios complexes faisant intervenir plusieurs phénomènes à la fois, comme l'alternance du chaud et du froid, la dessiccation et l'hydratation, le jour et la nuit, les processus lumineux etc.
Deux expériences ont été conduites pour confiner le métabolisme dans un liposome. Et dans les deux cas la nécessité de communiquer avec l'extérieur apparaît primordiale. Deamer [35] et al. (2002), expérimentent dans le modèle du «monde ARN» en encapsulant l'ARN dans des liposomes dont les chaines aliphatiques comportent 14 carbones pour que la membrane soit assez perméable pour laisser passer des nucléotides. Davis [36] et al. (2009) vont fabriquer une cellule artificielle minimale pouvant communiquer avec l'extérieur. Ils démontrent que l'encapsulation des ingrédients nécessaires à la réaction de formose peut produire des sucres dont des pentoses. Mais ils étaient obligés d'insérer dans la membranaire une protéine bactérienne, l'α-hémolysine, qui s'auto-assemble en un pore pour faire passer les sucres et communiquer ainsi avec l'extérieur.
Dans ce qui suit nous allons montrer que le métabolisme peut être initialisé à travers la bicouche lipidique grâce à des pores primitifs qui permettent d'interagir avec le milieu extérieur. Que ce métabolisme aura une partie spécifique, confinée à l'intérieur du liposome, qui correspond à la notion classique de métabolisme, et une partie dans la membrane qui fait office d'interface de communication. Pour cela nous allons d'abord présenter les expériences qui prouvent la synthèse abiotique des phospholipides dans la poche de pétrole.Synthèse abiotique des phospholipides et la formation des liposomes dans la poche de pétrole abiotique
- 1 - Esterification des acides gras :
-
- C'est une réaction athermique, un acide comme catalyseur; rendement maximum sur plusieurs mois sans catalyseur. ( Estérification )
- Conditions hydrothermales entre 100 et 300 °C. ( Rushdi [37] et al. 2006 )
- 2a.gras + glycerol ---> diacyl-glycérol (1,2/1,3 = 1)
- 7C, 2% en moyenne et jusqu'à 11% en présence de H2 (a. oxalique) et 100 °C ; plus de 34% de mono-acyl au-delà de 200 °C. Sur 72 h.
- a.gras + ethylene-glycol ---> acyl-ethylene-glycol 19C, 50% en moyenne jusqu'à 250 °C. Sur 72 h.
- Auto-assemblage en bicouche ( sans phosphate ni éthanolamine).
- 2 - Ethoxide : CH2=CH2 éthylène, synthétisé par FTT
-
- CH2 = CH2 + Cl2 + H2O ---> CH2Cl-OHCH2 + HCl éthlène chlorohydrine ( Weissermel [38] et al. 2003, p 146)
- 2( CH2Cl-OHCH2 ) + Ca(OH)2 ---> 2 (CH2CH2)O + CaCl2 + 2 H2O Ethoxide (même réactions que propène : voir glycerol)
- 3 - Ethanolamine
-
- (CH2CH2)O + NH3 ---> OH-CH2CH2-NH2 (Weissermel et al. 2003, p 159)
- solution 20-30% NH3 , 60-150 °C, 30-150 bar. (mono+di+tri) Ethanolamine
- HOCH2CHO + NH3 ---> (OH-CH2CH=NH) + H2O (imine, amination, glycolaldéhyde) A.D.Aubrey et al. (2009), Fig 5 [39]
- (OH-CH2CH=NH) + H2 ---> OH-CH2CH2-NH2 Ethanolamine
- conditions hydrothermales, 50-300 °C et 200 bars; 0.1 à 0.001M/kg de NH4HCO2.
- 4 - Esters phosphates
- 5 - Hydroformylation: ( Györgydeák [42] et al. 1998)
-
- CH2O + H2 + CO ---> Glycolaldéhyde, Glyceraldéhyde, sucres à 4 5 6 C.
- 140 °C, 120 bar, catalyseur: Co+triéthylamine
- 3 C2H5OH + NH3 ---> (C2H5)3N + 3 H2O alcool, triéthylamine
- 150-220 °C, 1 bars, catalyseur: Ni sur Si ou Al
- CO2 + 3H2 ---> CH3OH + H2O Méthanol
- 250 °C, 50-100 bar, catalyseur : Cu, ZnO, Al2O3. Ça ressemble à FTT.
- FTT produit des molécules à courte chaine quand la température est supérieure à l'optimale. McCollom [23] 1999 p.156.
- CH3OH ---> CH2O + H2 Méthanal
- 650 °C, 1 bar, catalyseur : Ag
- 3 CH3OH + K2Cr2O7 + 8 HClO4 ---> 3 CH2O + Cr2(ClO4)6 + 2 KClO4 + 7 H2O ( web [43] )
- à la place de HclO4 on peut utiliser H2SO4, solution 25 °C, 1 bar.
- 5 CH3OH + 2MnO4- + 6H+ ---> 5HCHO + 2Mn2+ + 8 H2O Oxydation d'un alcool
- solution 25 °C, 1 bar, milieu acide :H2SO4
- CH3CHO + HCN ---> CH3CH2OCN lactonitrile, catalyse basique, 1 bar, puis ( Weissermel et al. 2003, p 305)
- CH3CHO HCN + 2 H2O ---> CH3CHOHCO2H acide lactique, H2SO4, 100°C, 1 bar.
- 6 - Sucres phosphates: TMP, Trimétaphosphate
-
- a.Glycérique + TMP ---> 2-Glycérate-P + 3-Glycérate-P solution alcaline; ( Orgel [45] et al. 1996)
- NH3 + TMP ---> NH2-(PO3)3 ( Orgel [46] 2004 )
- Glycolaldéhyde + NH2-(PO3)3 ---> Glycolaldéhyde-P Mg++ pH 7 ( Eschenmoser [47] et al. 1999)
- R-CHOH-CHO + NH2-(PO3)3 ---> R-CHOPO3-CHO 27 aldéhyde-2-P ( Orgel [46] 2004 )
- 7 - Glycerol: CH2=CHCH3 propène, synthétisé par FTT
-
- CH2 = CHCH3 + Cl2 ---> CH2 = CHCH2Cl + HCl ( Weissermel [38] et al. 2003, p 296)
- à partir de 300 °C, 85% de produit à 500-510 °C, 1 bar ; chlorure d'allyle
- CH2 = CHCH2Cl + HOCl ---> CH2Cl-OHCHCH2Cl + CH2OH-ClCHCH2Cl (p 297)
- phase aqueuse, 25-30 °C, hypochlorate, dichlorohydroxypropane 70%, 30% respectivement
- 2 CH2Cl-OHCHCH2Cl + Ca(OH)2 ---> 2( CH2 =O= CHCH2Cl ) + CaCl2 + 2 H2O (p 297)
- 50-90 °C épichlorhydrine
- ( CH2 =O= CHCH2Cl ) + H2O ---> OHCH2-OHCH-CH2Cl (p 302)
- OHCH2-OHCH-CH2Cl + H2O ---> OHCH2-OHCH-CH2OH + HCl glycérol
- dans solution 10% soude caustique, 100-200 °C, haute pression.
- Glyceraldéhyde + H2 ---> Glycerol ( Hydrogenation )
- Ni, 60 bar. 50 °C ?
Références
- ↑ http://www.periodni.com/fr/p.html
- ↑ 2,0, 2,1 et 2,2 Broecker, W.S., and Peng, T.-H., 1982. Tracers in the Sea: Palisades, NY (Lamont-Doherty Geological Observatory).
- ↑ 3,0 et 3,1 Yamagata, Y., Watanabe, H., Saitoh, M. & Namba, T. (1991). Volcanic production of polyphosphates and its relevance to chemical evolution. Nature 352, 516-519.
- ↑ 4,0, 4,1, 4,2 et 4,3 G. Arrhenius, B. Sales, S. Mojzsis and T. Lee : Entropy and Charge in Molecular Evolution-the Case of Phosphate Journal of Theoretical Biology Volume 187, Issue 4, 21 August 1997, Pages 503-522
- ↑ http://webmineral.com/data/Brushite.shtml
- ↑ http://webmineral.com/data/Newberyite.shtml
- ↑ http://webmineral.com/data/Whitlockite.shtml
- ↑ A. Hall : Ammonium in granites and its petrogenetic significance . Earth-Science Reviews Volume 45, Issues 3-4, March 1999, Pages 145-165
- ↑ F.J.Stevenson :Chemical state of the nitrogen in rocks . Geochimica et Cosmochimica Acta, 1962, vol. 26, pp. 797-809
- ↑ Stuart R. Boyd, Anthony Hall and C.T. Pillinger : The measurement of δ15N in crustal rocks by static vacuum mass spectrometry: Application to the origin of the ammonium in the Cornubian batholith, southwest England. Geochimica et Cosmochimica Acta Volume 57, Issue 6, March 1993, Pages 1339-1347
- ↑ 11,0 et 11,1 Charlou J.L., Donval J.P., Fouquet Y., Jean-Baptiste P., Holm N., « Geochemistry of high H2 and CH4 vent fluids issuing from ultramafic rocks at the Rainbow hydrothermal field », Chemical Geology, vol. 191, 2002, p. 345-359.
- ↑ 12,0 et 12,1 Giora Proskurowski, Marvin D. Lilley, Jeffery S. Seewald, Gretchen L. Früh-Green, Eric J. Olson,1 John E. Lupton, Sean P. Sylva, Deborah S. Kelley: Abiogenic Hydrocarbon Production at Lost City Hydrothermal Field . Science vol 319, 1 février 2008 ;
- ↑ 13,0, 13,1 et 13,2 Bernard W. Evans : The Serpentinite Multisystem Revisited: Chrysotile is Metastable . International Geology Review, Vol. 46, 2004, p. 479–506 .
- ↑ http://webmineral.com/data/Chrysotile.shtml
- ↑ http://webmineral.com/data/Lizardite.shtml
- ↑ http://webmineral.com/data/Antigorite.shtml
- ↑ 17,0 et 17,1 Bernard W. Evans : Lizardite versus antigorite serpentinite: Magnetite, hydrogen, and life(?) . Geology, October 2010 v. 38; no. 10; p. 879–882; doi: 10.1130/G31158.1
- ↑ 18,0 et 18,1 T.M.McCollom et al. 2006:Carbon isotope composition of organic compounds produced by abiotic synthesis under hydrothermal conditions. Earth and Planetary Science Letters Volume 243, Issues 1-2, 15 March 2006, Pages 74-84
- ↑ http://www.annso.freesurf.fr/ProdH2.html
- ↑ T.M.McCollom et al. 2009:Thermodynamic constraints on hydrogen generation during serpentinization of ultramafic rocks. Geochimica et Cosmochimica Acta 73 (2009) 856–875
- ↑ N.G. Holm et al. 2006: Alkaline fluid circulation in ultramafic rocks and formation of nucleotide constituents: a hypothesis . Geochemical Transactions 2006, 7:7 doi:10.1186/1467-4866-7-7 .
- ↑ Thomas G. Kreutz, Eric D. Larson, Guangjian Liu, Robert H. Williams : Fischer-Tropsch Fuels from Coal and Biomass . 25th Annual International Pittsburgh Coal Conference 29 September–2 October, 2008 Pittsburgh, Pennsylvania, USA .
- ↑ 23,0, 23,1 et 23,2 T.M.McCollom et al. 1999: Lipid synthesis under hydrothermal conditions by fischer-tropsch-type reactions . Origins of Life and Evolution of the Biosphere 29: 153–166, 1999 .
- ↑ J.A.Brandes et al.1998: Abiotic nitrogen reduction on the early Earth. Nature vol 395 24 september 1998 .
- ↑ A. Smirnov :Abiotic ammonium formation in the presence of Ni-Fe metals and alloys and its implications for the Hadean nitrogen cycle. Geochemical Transactions 2008, 9:5 doi:10.1186/1467-4866-9-5;
- ↑ Henry P.Scott, Russell J.Hemley, Ho-kwang Mao, Dudley R.Herschbach, Laurence E.Fried, W.Michael Howard, and Sorin Bastea: Generation of methane in the Earth’s mantle: In situ high pressure–temperature measurements of carbonate reduction : PNAS September 28, 2004 vol. 101 no. 39 14023-14026
- ↑ . F. Kenney, Vladimir A. Kutcherov, Nikolai A. Bendeliani, and Vladimir A. Alekseev. The evolution of multicomponent systems at high pressures: VI. The thermodynamic stability of the hydrogen–carbon system: The genesis of hydrocarbons and the origin of petroleum. PNAS August 20, 2002, vol. 99 no. 17, 10976-10981
- ↑ 28,0 et 28,1 http://www.ifremer.fr/serpentine/fiches/fiche8.htm
- ↑ en.wikipedia.org/wiki/Pre-salt_layer
- ↑ Hu, Y. H. and Ruckenstein, E. (2006), Clathrate Hydrogen Hydrate—A Promising Material for Hydrogen Storage. Angewandte Chemie International Edition, 45:2011–2013. doi:10.1002/anie.200504149.
- ↑ T.A. Strobel et al. 2008 : clathrates of hydrogen with application towards hydrogen storage . Proceedings of the 6th International Conference on Gas Hydrates (ICGH 2008), Vancouver, British Columbia, CANADA, July 6-10, 2008.
- ↑ C.N.Murray et al. 1996 :permanent storage of carbon dioxide in the marine environment: the solid co2 penetrator. Energy Convers. Mgmt Vol. 37, Nos 6-8, pp. 1067-1072, 1996
- ↑ http://www.ac-nancy-metz.fr/enseign/physique/chim/Jumber/AMMONIAC/ammoniac_acide_nitrique.htm
- ↑ P.Avenier, M.Taoufik, A.Lesage, X.Solans-Monfort, A.Baudouin, A.de Mallmann, L.Veyre, J.-M.Basset, O.Eisenstein, L.Emsley and E.A.Quadrelli:Dinitrogen Dissociation on an Isolated Surface Tantalum Atom. Science 24 August 2007:Vol. 317 no. 5841 pp. 1056-1060 DOI: 10.1126/science.1143078. http://www.sciencemag.org/content/317/5841/1056.abstract
- ↑ Pierre-Alain Monnard, and David W. Deamer : Membrane Self-Assembly Processes: Steps Toward the First Cellular Life . The anatomical record, Volume 268, Issue 3, 1 November 2002, Pages: 196–207 . DOI 10.1002/ar.10154
- ↑ Paul M. Gardner, Klaus Winzer and Benjamin G. Davis : Sugar synthesis in a protocellular model leads to a cell signalling response in bacteria . Nature chemistry, vol 1, august 2009. DOI:10.1038/nchem.296
- ↑ Ahmed I. Rushdi, and Bernd R.T. Simoneit : Abiotic condensation synthesis of glyceride lipids and wax esters under simulated hydrothermal conditions . Origins of Life and Evolution of Biospheres (2006) 36: 93–108 . DOI: 10.1007/s11084-005-9001-6
- ↑ 38,0 et 38,1 Klaus Weissermel, Hans-Jürgen Arpe : Industrial organic chemistry. Wiley-VCH, 2003. pages 296-302.
- ↑ A. D. Aubrey & H. J. Cleaves & Jeffrey L. Bada: The Role of Submarine Hydrothermal Systems in the Synthesis of Amino Acids. Orig Life Evol Biosph (2009) 39:91–108 DOI 10.1007/s11084-008-9153-2
- ↑ http://orgprepdaily.wordpress.com/2007/02/10/ethanolamine-o-phosphate-and-n-palmitoylethanolamine-o-phosphate/
- ↑ James Jamieson Rae, Henry Davenport Kay, and Earl Judson King : Glycerophosphoric acid, the recognition and separation of α- and β-glycerophosphates. Biochem J. 1934; 28(1): 143–151.
- ↑ Zoltán Györgydeák,István F. Pelyvás. Monosaccharide sugars: chemical synthesis by chain elongation, degradation...(Page 8). Academic Press 1998.
- ↑ http://www.springerlink.com/content/j43707k803n57285/
- ↑ http://www.fsj.ualberta.ca/chimie/PowerPoint/aldéhyde_et_cétone.PPT
- ↑ Vera Kolb and Leslie E. Orgel . :Phosphorylation of glyceric acid in aqueous solution using trimetaphosphate . Origins of Life and Evolution of the Biosphere 26: 7-13, 1996.
- ↑ 46,0 et 46,1 Leslie E. Orgel : Prebiotic Chemistry and the Origin of the RNA World . Critical Reviews in Biochemistry and Molecular Biology, 39:99–123, 2004 . DOI: 10.1080/10409230490460765. (Fig.6 page 106).
- ↑ Ramanarayanan Krishnamurthy , Gustaf Arrhenius and Albert Eschenmoser : Formation of glycolaldehyde phosphate from glycolaldehyde in aqueous solution . Origins of Life and Evolution of the Biosphere 29: 333–354, 1999.
1 commentaire
-
Par mekali le 8 Septembre 2012 à 14:59
début de rédaction dans wikiversité 050112
Sommaire
- 1 Introduction
- 2 Chiralité de la serine prébiotique
- 3 Chiralité du glycérol-P prébiotique
- 4 Les caractéristiques physico-chimiques des acide aminés prébiotiques
- 5 De l'homochiralité prébiotique à l'homochiralité biotique
- 5.1 Les processus prébiotiques pour la mise en place de l'homochiralité
- 5.2 Hypothèses sur l'évolution moléculaire du processus prébiotique au métabolisme biotique
- 5.2.1 Introduction
- 5.2.2 Au-delà de la chiralité il y a la conformation spatiale en générale
- 5.2.3 La cohésion mécanique du liposome est à l'origine de la chiralité et de l'homochiralité des sucres
- 5.2.4 La CMP positionne le tétraèdre du phosphate pour estérifier le bras libre dans la bonne configuration nécessaire à la cohésion mécanique du liposome.
- 5.2.5 La cohésion mécanique du liposome et l'initialisation du métabolisme prébiotique
- 6 Références
De l'origine mécanique et géométrique de la chiralité et de l'homochiralité du glycérol-phosphate, du glycéraldéhyde-P et des acides aminés prébiotiques ainsi que des principales caractéristiques physico-chimiques de ces acides aminés.
Résumé
Le rapprochement des phospholipides, dans la bicouche d'un liposome, provoque leur rotation autour de leur axe d'acides gras, générant une force qui rapproche les 2 feuillets de la bicouche. Dans cette étude théorique je montre que pour obtenir la plus grande cohésion du liposome, par ces forces, la sérine de la tête hydrophile doit avoir une chiralité L. Dans le cas où la tête hydrophile serait absente les acides aminés de chiralité L pourraient contribuer à cette cohésion en prenant la place de la L-sérine. Certains coenzymes présentant une configuration analogue à l'éthanolamine pourraient y contribuer aussi. C'est le cas de la pyridoxamine, de la thiamine et du tétrahydrofolate prébiotiques.
Le regroupement des acides aminés de chiralité L et de la pyridoxamine sur la paroi pourrait initialiser le métabolisme prébiotique de ces seuls acides aminés. C'est ce qui expliquerait l'origine de l'homochiralté des acides aminés dans le vivant.
Par ailleurs je montre que, dans la tête hydrophile, l'estérification du glycérol-phosphate à 2 acides gras passe par le positionnement de la dihydroxyacétone-phosphate et du L-glycéraldéhyde-3-phosphate , mais pas du D-glycéraldéhyde-3-phosphate, avant leur hydrogénation en glycérol-3-phosphate. L'accumulation du D-glycéraldéhyde-3-phosphate dans le cytoplasme déplacerait les équilibres thermodynamiques vers la synthèse de la D-dATP à partir du D-glycéraldéhyde-3-phosphate, de l'acétaldéhyde et de l'adénine prébiotique, réaction qui ne nécessite pas de coenzyme dans le métabolisme biotique. La D-dATP et la thiamine, plus le métabolisme prébiotique des acides aminés L sur la paroi, initialiseraient les voies des D-pentose-phosphates et des D-nucléotides à partir de la réaction D-glycéraldéhyde-3-phosphate + dihydroxyacétone-phosphate et des bases nucléiques prébiotiques.
L'épuisement du glycéraldéhyde prébiotique (racémique) et le métabolisme biotique naissant, dominé par le D-glycéraldéhyde-3-phosphate, expliqueraient l'origine de l'homochiralité des sucres dans le vivant.1. Introduction
17/11/11
Dans ce qui suit j'ai abrégé les termes suivants parce qu'ils sont très utilisés:
acide aminé: aa
acide gras: ag
phospholipide: PLD
phosphate: P.
Cette étude théorique est faite dans le cadre de l'hypothèse d'une évolution moléculaire prébiotique qui se déroulerait dans une poche de pétrole abiotique d'origine géochimique ( pétrole prébiotique, en préparation). Le développement théorique et bibliographique de cette hypothèse a montré qu'il était tout à fait possible que les liposomes puissent se former dans ces conditions à partir de vésicules aqueuses présentes dans la phase huile et qui migreraient dans la phase eau pour former ces liposomes. La paroi des vésicules seraient formées par la tête carboxylique des acides gras et le contenu en eau de ces vésicules, ainsi que la phase eau principale, contiendraient les petites molécules hydrophiles dont le glycérol, l'éthanolamine, le phosphate et certains acides aminés racémiques, dont la sérine, pour synthétiser par estérification la tête hydrophile que l'on connait.
C'est en essayant d'imaginer l'agencement de ces petites molécules en tête hydrophile (dans la vésicule ou le liposome) et la fixation de cette tête sur les ag, qu'il m'est apparu que ce processus puisse imposer la chiralité du glycérol-P et des aa ainsi que la séquestration de ces aa sur les parois, en fonction de leur chiralité, mais aussi de leur longueur, de leur volume et de la fonction chimique portée par leur radical.
Dans un premier temps nous étudierons l'enchainement des molécules de PLD, les unes aux autres, grâce à l'éthanolamine ou à la choline ou à la sérine, qui font partie intégrante de la tête hydrophile. Nous montrerons que cet agencement par enchainement est à l'origine de la chiralité de la sérine. Nous étudierons ensuite la chiralité du glycérol-P dont l'origine est à trouver dans un compromis entre l'encombrement stérique et la répulsion électronique entre la tête carboxylique de l'ag proche du P et ce dernier.
Un chapitre particulier sera dédié à la séquestration des aa libres parce qu'ils vont se mettre à la place des têtes hydrophiles, grâce à leur amine, lors de l'agencement dans les vésicules ou lors de la perte de ces têtes en réaction au milieu extérieur.
Nous terminerons enfin par les propriétés de cette homochiralité de la paroi liposomale qui peut être à l'origine de l'initialisation du métabolisme prébiotique dans la bicouche même, hypothèse émise dans le travail sur la chimio-osmose prébiotique. L'homochiralité du métabolisme que nous connaisons serait originaire de celle de la bicouche prébiotique.2. Chiralité de la serine prébiotique
La serine est attachée par liaison ester au phosphate comme le sont l'éthanolamine, la choline et le glycérol. Cette partie de la tête hydrophile constitue un bras mobile. La 2ème partie de la tête hydrophile est le bras fixe, constitué du glycérol-P fixé à 2 ag. Même quand la molécule de PLD est isolée des autres molécules de PLD, le bras fixe présente une grande inertie. Et lorsque 2 molécules de PLD se rapprochent par la force de la liaison hydrogène (NH2) ou ionique (NH3+), vers les 2 oxygènes libres du P, et par la force de l'hydrophobicité (rapprochement des 4 ag), c'est le bras libre qui bougera en premier. Et ceci d'autant plus vite et plus fort que la liaison hydrogène (ou ionique) agit à plus grande distance que ne le font les forces de Van der Walls pour l'hydrophobicité des ag. Ceci entraine obligatoirement une rotation de la molécule de PLD entière, autour des 2 ag, car l'amine NH2 (NH3+) n'est pas alignée avec le bras libre, et le P dont elle se rapproche est massif et très proche des ag.
Si l'on se positionne au-dessus du PLD avec les ag en dessous du plan défini par 3 oxygènes du P, une rotation anti-horaire soulèverait la molécule de PLD et une rotation horaire l'enfoncerait. La rotation horaire rapproche donc le feuillet en question vers l'autre feuillet de la bicouche, ou vers la phase huile pour la vésicule aqueuse. La rotation horaire est donc un facteur de cohésion très important si l'on tient compte du grand nombre de molécules de PLD que contient la bicouche. Et c'est cet agencement qui déterminera la chiralité de la serine et de la position de l'amine NH2 (NH3+) de l'éthanolamine.2.1 Positionnement du NH2 de l'éthanolamine dans la molécule de phospholipide pour obtenir la plus grande cohésion possible du liposome
Si on regarde de face les 2 oxygènes libres du phosphate (voir dessin 1), NH2 peut occuper 4 positions possibles: avant-gauche(VG), arrière-gauche(RG), avant-droite(VD), arrière-droite(RD). La position RG est la seule à donner un enchainement, avec rotation horaire, d'un grand nombre de PLD.
En effet, les 2 positions VG et RD tournent dans le sens anti-horaire pour rejoindre le P. Les 2 autres positions tournent dans le sens horaire, mais la position VD ne peut s'agencer qu'avec 1 seul autre PLD, car NH2 et les 2 oxygènes du P sont du même côté et se neutralisent mutuellement. Par contre la position RG permet d'agencer à la queue leu-leu et de façon illimitée les PLD les uns aux autres tout en assurant le rapprochement des 2 feuillets du liposome par rotation horaire.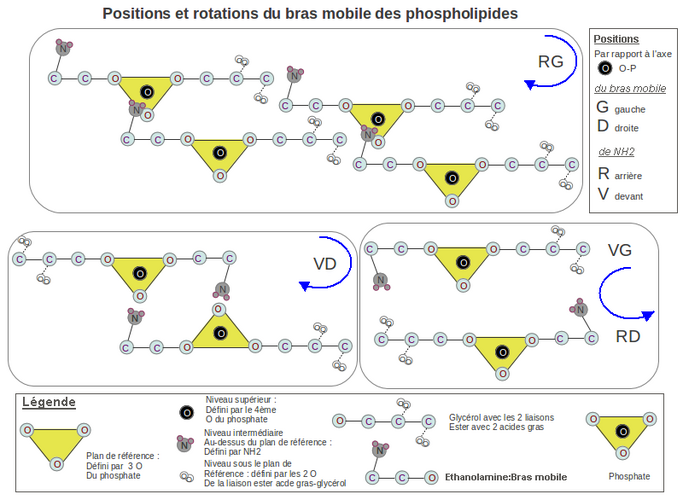
Dessin 1. Hypothèse de l'origine mécanique de la chiralité dans les phospholipides. Ici est traité le positionnement de l’éthanolamine par rapport au phosphate où son déplacement vers lui pour établir une liaison hydrogène entraîne une rotation. Cette hypothèse stipule que la rotation horaire et le positionnement à la queue leu-leu des molécules phospholipides (encadré RG) entraîne le rapprochement des 2 feuillets du liposome, augmentant sa cohésion. Dans cette configuration la sérine aurait une configuration L de type lévogyre, car son carboxyle ne peut se mettre sous le plan de référence défini par le phosphate (voir la légende dans le dessin) et serait alors à proximité des carboxyles des 2 acides gras des molécules voisines.
2.2 La chiralité de la sérine est de type lévogyre (L).
Nous venons de voir l'origine mécanique du positionnement du NH2 de l'éthanolamine. Ceci est valable aussi pour le NH2 de la sérine. Le carbone de la fonction carboxylique de la sérine va se positionner, lui, par répulsion électronique. En effet si on dessine l'agencement de plusieurs PLD voisins (en vue aérienne, voir dessin 1-RG), on voit que les PLD d'une rangée s'emboitent dans la rangée voisine et que le carbone de l'amine (NH2) se trouve au même niveau et aligné le long d'une oblique, avec les carbones portant les esters des glycérols.
Si on positionne la fonction carboxylique de la sérine sous le plan défini par le P (ce qui donnerait la D-sérine), alors elle se trouvera entourée des 4 oxygènes des 2 liaisons ester appartenant aux 2 molécules de PLD successives. Il y aura alors, dans un espace exigu, 6 oxygènes pour un seul hydrogène pour faire la liaison, dans le meilleurs des cas suivant le pH. Comme les 2 fonctions carboxyliques des 2 ag sont fixes, suite à la cohésion globale du liposome, celle de la sérine, mobile, est expulsé automatiquement par répulsion électronique au-dessus du plan défini par le P. Nous avons alors de la L-sérine.
Dans le cas des archées ce n'est plus la répulsion électronique qui intervient, mais l'encombrement stérique. Le carbone de la fonction carboxylique de la D-sérine s'intercalerait entre les têtes des ag, ce qui écarte les 2 PLD de la rangée voisine, augmentant fortement l'encombrement. Or l'encombrement stérique doit être minimal pour ces procaryotes, comme on le verra aussi pour la chiralité de leur glycérol-P du bras fixe.3. Chiralité du glycérol-P prébiotique
La chiralité du glycérol-P doit être indépendante de la position du bras mobile comme c'est le cas entre bactéries et archées. Ces lignées assurent la cohésion du liposome avec le même principe mécanique qu'on vient de voir, mais les chiralités de leur glycérol-P du bras fixe sont opposées. La chiralité du bras fixe dépend de la liaison du glycérol aux chaines aliphatiques. En effet la liaison ester est volumineuse et répulse les électrons avec ses 2 oxygènes, alors que la liaison éther n'a qu'un oxygène et est moins répulsive que chacun des 2 oxygènes de la liaison ester (la liaison éther est connue pour être très stable, étant très peu réactive).
Chez les archées, avec la liaison éther sur les ag pour minimiser l'encombrement stérique, les 2 liaisons éther peuvent se mettre toutes les 2 du même côté que les 2 oxygènes libres du P, pour remplir l'espace vacant sous le demi-plan de référence défini par le P (voir Dessin 2). Comme la position de la sérine par rapport au P est imposée mécaniquement, comme on l'a vue précédemment, son amine (NH2) et l'oxygène de la première liaison éther se trouvent, du coup, chacun dans un demi-plan différent, la chiralité du glycérol-P est alors de type lévogyre comme la sérine, puisqu'ils se superposent par rotation. C'est un L-glycérol-3P si l'alcool terminal se trouvait au-dessus des carbones du glycérol comme le CO2H de la sérine se trouve au-dessus de la membrane. Or pour diminuer l'encombrement stérique l'alcool terminal du glycérol doit être au-dessous de ses carbones. C'est alors un D-glycérol-3P ou L-glycérol-1P (sn-glycérol-1P), celui qu'on trouve chez les archées.
Avec la liaison ester, la répulsion électronique entre elle et les oxygènes libres du P, positionne automatiquement la première chaîne aliphatique la plus proche du P, du même côté que l'amine de la sérine, de l'autre côté du demi-plan défini par le P (voir Dessin 1-RG). Et par encombrement et répulsion avec la première, la deuxième chaîne aliphatique se trouve du côté du demi-plan contenant le P et sous les carbones du glycérol pour les mêmes raisons d'encombrement que pour les archées. D'où le L-glycérol-3P (sn-glycérol-3P) des bactéries ou D-glycérol-1P (voir KEGG pour les synonymes ).
Il faut aussi signaler que dans la vésicule ou sur le feuillet interne du liposome, lors de la mise en place des têtes hydrophiles, les ag sont collés côte à côte et, l'hydroxyle de l'un faisant face à l'oxygène de l'autre, la liaison hydrogène entre eux se trouve renforcée. Lors de la fixation des têtes hydrophiles sur un champs de carbones carboxyliques, la moitié du travail est faite et reste juste à positionner la tête.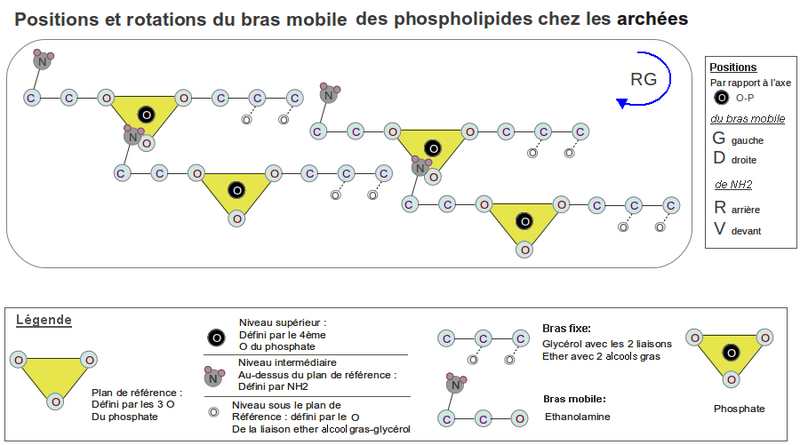
Dessin 2. Voir Dessin 1 pour la légende.
4. Les caractéristiques physico-chimiques des acide aminés prébiotiques
4.1 Les acides aminés dans la théorie de la chimio-osmose prébiotique
J'ai étudié déjà l’interaction aa-liposome dans l'article théorique sur la chimio-osmose prébiotique. J'ai supposé alors que pour former les pores prébiotiques, qui, à l'instar des ionophores produits par certains procaryotes, contiennent des acides aminés (D et L) et des acides alpha-hydroxylés (D et L), les aa devaient s'intercaler entre le P et l'amine, avec liaison hydrogène des 2 côtés. Leur radical se positionnant du coup verticalement à la bicouche, soit dans la bicouche, soit à l'extérieur.
Le passage des aa à travers la bicouche, par des pores prébiotiques ou unitairement, m'a conduit à supposer que le métabolisme pourrait bien commencer dans la bicouche qui leur sert alors d’échafaudage pour les positionner les uns à côté des autres. L'efficacité de ces groupements d'aa serait de plus en plus grande si leur homochiralité augmente de plus en plus. Mais je n'avais émis aucune hypothèse sur le type, D ou L, de la chiralité finale de ces aa.
De même, la ressemblance de 13 aa biotiques avec l'éthanolamine et la sérine pour la longueur de leur chaine carbonée (2 C) entre la tête et la fonction portée par le radical, ainsi que les contraintes qu'ils doivent subir lors de leur passage à travers la membrane, me laissaient deviner que les aa prébiotiques devaient être courts et hydrophobes. Par contre les radicaux ionisés ou très réactifs posaient problème pour le passage à travers la membrane. Ce sont les radicaux ionisables: acides, amides, amines ainsi que ceux de l'histidine et de l'arginine.
La séquestration des aa dans la théorie de la cohésion mécanique du liposome, que nous allons voir juste après explique leur origine chirale L, origine que n'arrive pas à introduire la théorie de la chimio-osmose prébiotique. Cependant les 2 processus, diffusion à travers la membrane et séquestration des aa que nous allons voir tout de suite, ne sont pas exclusifs l'un l'autre et peuvent coexister en même temps.4.2 La séquestration des acides aminés dans la théorie de la cohésion mécanique du liposome
4.2.1 Principe:
Nous avons vu précédemment que la chiralité de la sérine dans le liposome a son origine dans une force qui rapproche les 2 feuillets. C'est une force perpendiculaire à la membrane. Mais elle n'est possible que si la sérine est rattachée au P du bras fixe par une liaison ester. D'ailleurs toute amine rattachée au P peut servir cette cohésion. C'est le cas de l'éthanolamine, de la choline mais aussi du glycérol avec son hydroxyl terminal pour la liaison hydrogène.
Cependant la cohésion mécanique du liposome a 2 composantes, la composante perpendiculaire à la surface du liposome que nous venons de voir et que je nommerai verticale, mais surtout une 2ème composante perpendiculaire au plan défini par les 2 ag du PLD et que je nommerai horizontale, plus forte, et dont le sens va de l'amine d'un PLD au P du PLD suivant. Il n'y a pas de force de cohésion perpendiculaire à la force horizontale et contenue dans le plan des ag d'un même PLD. C'est ce qui autorise les glissements des chaines de PLD les unes par rapport aux autres et différencie le liposome de la rigidité du cristal.
La cohésion horizontale a 2 composantes parallèles, la première, la liaison hydrogène, amine-P, entre 2 têtes hydrophiles consécutives et la deuxième, constituée par les liaisons de Van der Walls entre les queues aliphatiques des 2 PLD consécutifs. Si ces dernières constituent le fondement même de la formation des vésicules, puisqu'in vitro on peut fabriquer des vésicules avec des ag seulement, les liaisons hydrogènes de la première composante peuvent se faire et se défaire rapidement localement, tout en maintenant la cohésion globale horizontale. Ce qui compense la grande inertie des queues hydrophobes. Dans les vésicules sans bras mobile,avec des ag seulement, les têtes carboxyliques disposées en tête-bêche peuvent former des liaisons hydrogènes, mais ne forment pas une cohésion globale horizontale.
Aussi, s'il arrive que la liaison ester du bras mobile vienne à être hydrolysée, ou que toute la tête hydrophile vienne à manquer, la place est libre pour toute autre molécule portant une amine. C'est ce que j'appelle la séquestration.4.2.2 La chiralité des acides aminés séquestrés:
Dans le cas des aa séquestrés, la première composante de la cohésion horizontale n'est rétablie entièrement que si le NH2 (ou même un OH) est accompagné d'un P ou d'un carbone carboxylique, comme c'est le cas des aa, pour rattacher l'amine du PLD de l'arrière et qui se trouve à son niveau. Ces aa vont être bien sûr de type lévogyre (L) comme la sérine, puisqu'on se trouve toujours dans la même configuration où il n'y a pas de brèche dans le liposome (trou sans ag) et que les chaînes de PLD se côtoient (voir Dessin 1-RG ci-dessus).
4.2.3 Les autres caractéristiques physico-chimiques des acides aminés séquestrés:
L'espace libéré par la tête hydrophile a une surface bien définie. Ça serait là l'origine géométrique de ces aa prébiotiques. Mais leur origine est aussi mécanique puisqu'ils agissent mécaniquement comme la sérine liée au P. Ils ont donc une origine mécanique et géométrique.
- Horizontalité: Ces aa sont disposés horizontalement à l'inverse de ceux qui traversent la membrane dont la disposition pourrait être aléatoire.
- La longueur maximale de ces aa est approximativement équivalente à 7 atomes (voir Dessin 1 ci-dessus), sans tenir compte du carbone carboxylique. Et l'aa ne peut dépasser cette longueur, sinon il empêcherait le glissement des chaînes de PLD voisines et vice-versa.
- La largeur maximum de ces aa, ou épaisseur, est limitée à 4 carbones (voir Dessin 1 ci-dessus). Ceci conduit à de petites chaînes latérales comme avec val, thr, leu, ile, arg, ou à l'épaisseur d'un seul cycle aromatique comme phe, trp, tyr, his.
- En hauteur, c'est-à-dire en dehors de la membrane, il n'y a pas de limite.
4.2.4 Les fonctions chimiques portées par le radical des acides aminés séquestrés:
Il n'est pas question ici de retrouver les chaînes des réactions chimiques du réseau métabolique prébiotique. Mais si la séquestration par la cohésion mécanique favorise certains aa et en exclut d'autres, cela nous permettrait d'entrevoir l'initialisation du métabolisme prébiotique à partir de certains groupements d'aa.
- Les acides carboxyliques et les amides:
Il est remarquable de constater qu'il y ait 2 fonctions carboxyliques et 2 fonctions amides courtes à 1 et 2 atomes de plus que la sérine, en longueur, chez les aa biotiques! Pourquoi ces longueurs et ce double emploi unique parmi les 20 aa biotiques? La réponse peut-elle provenir de la cohésion mécanique? On peut effectivement émettre l'hypothèse qu'à ces longueurs ces 2 fonctions se trouvent au niveau du P de la tête perdue et qu'elles puissent attirer l'amine du PLD de l'arrière par liaison hydrogène. Ce qui rétablirait presque entièrement la première composante de la cohésion horizontale comme le ferait un P, alors que la fonction carboxylique de la tête des autres aa, décalée par rapport à l'amine du PLD arrière, ajouterait un déséquilibre à ce rétablissement.
On peut aussi supposer, dans le cas où la tête ne garde que le glycérol du bras fixe, que l'hydroxyle libre de celui-ci puisse établir une liaison hydrogène avec ces fonctions et une liaison ester même avec la fonction carboxylique. Ce qui rétablirait et la première composante horizontale et la composante verticale de la cohésion mécanique.
Par contre une fonction carboxylique ou amide d'un aa plus long arriverait au-dessus des ag, serait repoussée par répulsion électronique et destabiliserait la fixation de l'aa.- Les radicaux portant un hydroxyle ou une amine et arrivant au niveau des ag, peuvent établir des liaisons hydrogène avec ces derniers. Ce qui rétablirait partiellement la composante verticale de la cohésion. C'est le cas de arg et tyr (4 atomes de plus que la sérine en longueur), de lys et his (plus 3 atomes).
- Les radicaux hydrophobes, peu réactifs devraient remplir toute la largeur de l'espace libéré. C'est une consolidation par encombrement stérique. C'est le cas de 9 aa biotiques qui sont soit branchés (leu, ile, val, thr), soit aromatiques(phe, trp, pro), soit contenant un atome de soufre(met, cys) qui a autant d'influence stérique par son volume et son noyau que l'atome de phosphore.
- Il n'y a que 2 aa biotiques qui n'apportent que leur tête pour la cohésion mécanique: ala, gly.
- Les problèmes que soulèvent certains aa biotiques tels que met, cys, pro, trp:
Tout radical portant un atome à forte action sur son voisinage devrait être exclu, comme les métaux de transitions, les halogènes, les cations mais aussi le soufre. Dans ce cas la cys peut avoir une origine non mécanique et provenir de la thiolyse de la liaison ester sérine-P, par exemple. Reste l'origine de la met. La pro peut-elle établir une liaison hydrogène sous sa forme cyclique? Ou bien la pro prébiotique était-elle linéaire? Le trp devrait poser un problème de hauteur puisqu'aucun aa biotique n'a de la hauteur! Est-ce que la citruline et l'ornitine feraient parties des aa prébiotiques séquestrés?
5. De l'homochiralité prébiotique à l'homochiralité biotique
Nous venons de voir les principes théoriques de la mise en place de l'homochiralité, en supposant que les molécules nécessaires sont, soit préexistantes en quantité dans le milieu réactionnel (prébiotique) comme les ag, soit qu'elles puissent être récupérées facilement par déplacement des équilibres thermodynamiques.
Nous allons étudier dans ce chapitre le rapprochement de ces 2 processus en tenant compte des conditions plus contraignantes de l'hypothèse de la poche de pétrole prébiotique et en prenant des circuits métaboliques biotiques les moins complexes possibles. Et cela pour essayer d'imaginer l'évolution moléculaire qui pourrait s'opérer du prébiotique au biotique pour cette mise en place de l'homochiralité.5.1 Les processus prébiotiques pour la mise en place de l'homochiralité
Ce sont d'abord les processus fondamentaux de la thermodynamique statistique en milieu liquide (huile ou eau). Les molécules diffusent librement dans le milieu, et interagissent entre elles avec des énergies augmentant avec la température. Dans l'environnement organisé d'une cellule, les petites molécules ne sont pas libres et sont contrôlées et transportées même par des macromolécules. Le cytoplasme est considéré comme un "gel colloïdal".
C'est cette différence fondamentale qui nous interdit de plaquer les réactions enzymatiques du métabolisme biotiques aux réactions chimiques d'un mélange complexe, liquide, sans macromolécules. Le point de départ de l'évolution moléculaire, dans notre hypothèse de poche de pétrole prébiotique, étant l'auto-assemblage des ag synthétisés abiotiquement, notre objectif que nous avons introduit ci-dessus, c'est de mettre en parallèle ces 2 mondes, de les rapprocher en prenant les réactions du métabolisme biotique proches des réactions de la chimie organique, et de considérer les conditions géochimiques de la poche de pétrole les plus proches des caractéristiques physico-chimiques du vivant.5.1.1 Les conditions de la poche de pétrole prébiotique:
Les conditions physico-chimiques les plus proches des caractéristiques du vivant sont celles, par exemple, des bactéries vivants dans les poches de pétrole "fossile", à 55 °C et sous 400-800 bars [1]. Mais on peut s'en éloigner encore un peu pour dépasser légèrement la température maximale de la multiplication des extrêmophiles de 113 °C, à 150 °C; la pression a moins d'influence sur les bactéries et on peut la doubler pour atteindre 1,5 kbar comme dans le gisement de pétrole de Tupi au large du Brésil [2].
Je pense que dans ces conditions, les réactions qui s'y déroulent peuvent être en équilibre thermodynamique et donc produire en continu, certaines molécules, si celles-ci sont piégées par ailleurs. C'est le cas de l'hydroformylation qui produit le glycéraldéhyde et la dihydroxyacétone (100-140 °C, 120 bars, à partir de CH2O H2 CO; Györgydeák [3] et al. 1998), de l'hydrogénation du glycéraldéhyde en glycérol (50 °C, 60 bars, en présence de H2) et la synthèse d'aa et d'éthanolamine (A.D.Aubrey et al. (2009), Fig 5 [4]) et de la choline (synthèse analogue à la triméthylamine).
Dans l'hypothèse d'une formation du pétrole abiotique par diagénèse (voir pétrole prébiotique), où pression et température s'élèvent progressivement, CH2O H2 CO NH3, issus des clathrates de départ (J.L Charlou, [5] Ifremer: Serpentinisation et synthèse inorganique d'hydrogène, méthane et hydrocarbures le long de la dorsale médio Atlantique), pourraient exister et former glycérol, glycéraldéhyde, dihydroxyacétone, éthanolamine, choline ainsi que quelques aa en petites quantités.
Dans l'hypothèse d'une formation brutale du pétrole abiotique (voir pétrole prébiotique) par métamorphisme, lors de la subduction ou proche de ces zones (cônes d'accrétion), les conditions de pression et de température sont au niveau supérieur des limites pour la synthèse du pétrole par le processus FTT. Ces conditions rendraient possible la synthèse du glycérol et de l'éthanolamine directement comme dans les procédés industriels (500 °C et hautes pressions) à partir des molécules d'éthylène et de propène issues du processus FTT. Ces hautes températures ne permettraient pas la formation des têtes hydrophiles. Il faudrait attendre la migration des poches de pétroles vers des zones à températures plus modérées. A ces températures la synthèse de ces 2 molécules, éthylène et propène, ne se faisant plus, il faudrait supposer que de grandes quantités de glycérol et d'éthanolamine aient été synthétisées auparavant, en même temps que les ag et dans les mêmes concentrations, pour subvenir à la formation des têtes hydrophiles. Aucun résultat, dans le domaine du pétrole "fossile" ou synthétique n'a, jusqu'à présent été rapporté dans ce sens, pour le glycérol (pétrole fossile et synthétique, abscence de N2) ou pour l'éthanolamine (pétrole fossile, présence de N2).5.1.2 La mise en place de l'homochiralité prébiotique:
Plaçons-nous donc dans le cas d'une poche de pétrole prébiotique formée lentement par diagénèse, où un réseau de réactions chimiques existe en équilibre thermodynamique et qui peut fournir en continu P, glycérol, GA, DHA, éthanolamine, choline, sérine et d'autres aa. Les concentrations de ces molécules n'ont pas d'importance, parce que seul compte la possibilité de piégeage qui déplace les équilibres. Et ce piégeage se fait par les ag et les PLD intégrés dans des macro-structures cohérentes, difficiles à hydrolyser, comme les vésicules dans la phase huile ou les liposomes dans la phase eau.
Cependant pour la fixation des têtes hydrophiles sur les têtes des ag, toutes les réactions sont des estérifications. Or dans l'eau l'hydrolyse tend à les défaire. Et la phase huile, qui permettrait de les conserver, est réduite à l'épaisseur de la bicouche, si cette fixation devait se faire dans des bicouches d'ag de la phase eau (équivalentes des liposomes sans tête hydrophile).- Les vésicules de la phase huile.
Dans la phase huile les vésicules concentrent toutes les molécules hydrophiles avec en principe peu d'eau. Nous avons vu dans l'article de la chimio-osmose prébiotique, que le liposome dans la phase eau représente le fondement même du vivant, puisqu'il délimite un intérieur spécifique, d'un extérieur changeant et qu'il permet la croissance de l'organisation à l'intérieur par une communication (électronique et ionique) à travers la membrane. Les vésicules de la phase huile, elles, ne peuvent pas communiquer avec l'extérieur, l'huile constituant un mur infiniment épais, ne permettant que l'arrivée des molécules hydrophiles. Les vésicules sont vites isolées les unes des autres.
Par contre les vésicules de la phase huile possèdent une vraie interface eau/huile. Dans une telle interface, les molécules de chimie organique produites pendant le processus de la formation de la poche de pétrole prébiotique, peuvent occuper les 2 phases et créeraient même une phase intermédiaire pour les molécules qui sont plus ou moins solubles dans les 2 premières phases. Cette zone intermédiaire peut être très mince, mais elle permet, avec les 2 autres phases, toutes les réactions possibles suivant cette graduation de la solvatation. Et notamment elle permet l'estérification des têtes des ag par les têtes hydrophiles, l'hydrolyse des liaisons ésters devenant de plus en plus difficile avec l'étendue de la surface phospholipidique.- Synthèse et fixation par estérification de la tête hydrophile.
D'après ce que nous savons, l'estérification est une réaction réversible; le sens du déplacement de l'équilibre dépend uniquement du solvant; pour un solvant donné l'état d'équilibre ne dépend ni de la température ni de la pression; les réactions sont très lentes en l’absence de catalyseur; le catalyseur est le proton H+ issu de HCl, H2SO4, H3PO4; les têtes carboxyliques sont aussi catalyseurs (auto-catalyse): voir les esters dans wikipédia.
Aussi nous allons supposer que les estérifications qui nous concernent sont possibles dans l'environnement de la poche de pétrole prébiotique et qu'elles se réalisent lentement, et même très lentement étant donné que le temps géologique n'est pas déterminé. Cependant par comparaison avec le métabolisme biotique, nous allons voir dans ce qui suit, que le rôle de celui-ci va agir surtout sur le nombre d'intermédiaires du réseaux des réactions chimiques.
Par exemple le déplacement d'équilibre du mélange racémique du glycérol-P vers le glycérol-3P fixé par les ag, va nécessiter l'enlèvement du glycérol-1P ou du 2P de la surface des ag, leur hydrolyse, puis l'estérification du glycérol libéré en glycérol-3P, puis diffusion de ce dernier vers la surface d'ag. La fixation de ce dernier déplacera alors effectivement l'équilibre du GA en glycérol (réaction d'hydrogénation).
Devant ce grand nombre d'intermédiaires et le potentiel catalytique de la surface des ag et/ou des têtes hydrophiles, il est tout à fait légitime de considérer l'hydrogénation du glycéraldéhyde-P racémique (ou de la dihydroxyacétone) en glycérol-3P après fixation du 1er sur l'ag. Car la tautomérie céto-énolique de la fonction aldéhyde leur permet d'adopter la conformation imposée par la surface, comme nous l'avons vu dans l'approche théorique de la chiralité du liposome (Dessin 1-RG, et Dessin 2). Une fois le glycéraldéhyde-P fixé et la conformation glycéraldéhyde-3P réalisée, alors l'hydrogénation rendue possible par la présence de H2, comme nous l'avons suggérer ci-dessus (Les conditions de la poche de pétrole prébiotique), et par l'influence catalytique de la surface, peut se faire facilement.
Chez le vivant l'inertie engendrée par les déplacements de ces nombreux équilibres est éliminée grâce au transport des petites molécules par les macromolécules, et seule la conformation adéquate est synthétisée et protégée de la racémisation par ces macromolécules. Par ailleurs on conçoit facilement que la mobilité de certaines enzymes dans la phase eau (l'intérieur du liposome ) sera une des 1ères étapes de l'évolution moléculaire, car elles pourront agir successivement à plusieurs endroits de la surface, en emmenant les petites molécules nécessaires avec elles. Sinon, sans cette mobilité, les petites molécules se fixeraient au hasard, puis suivant l'environnement surfacique, la catalyse pourrait être plus ou moins efficace, et les déplacements d'équilibre, en cas d'erreur, rallongeraient énormément la réalisation.- Formation des liposomes.
Nous avons vu précédemment que les vésicules bicouches d'ag dans l'eau ne conviennent pas pour la formation des molécules de phospholipides. De même la formation des liposomes hétérogènes, formés d'assemblage au hasard de phospholipides et d'ag, comme décrit dans la littérature ( Segré 2000 [6], par exemple ), devraient aboutir difficilement, pour les mêmes raisons, à un liposome homogène, ou structuré (têtes sérine, choline et éthanolamine) avec très peu d'ag nus.
Le scénario de la formation des liposomes que je décris ici, suppose que les conditions de température et de pression sont stables sur une longue période, que la poche de pétrole n'est pas soumise, pendant cette période, à des turbulances, permettant ainsi la formation de phospholipides dans les vésicules la phase huile. L'interface eau/huile des 2 phases globales sera, dans cette hypothèse, constituée, avec la durée, de phospholipides provenant de vésicules ayant migré vers la phase globale eau et y ayant versé leur eau.
Pour qu'une vésicule PLD puisse donner une bicouche PLD, il faut que son feuillet lipidique ne se rompt pas et qu'elle ne rompt pas le feuillet PLD constituant l'interface de la phase globale. Ces vésicules vont s'accumuler en bas de la phase huile, et quand la pression de celles qui sont au-dessus devient assez grande, celles qui sont en contact avec le feuillet global, vont soit éclater comme nous l'avons décrit ci-dessus, soit se détacher pour entrer dans la phase eau globale en s'entourant d'un morceau du feuillet global d'une longueur équivalente à la longueur de son feuillet interne.
Ce scénario de la formation des liposomes donne 2 feuillets homogènes mais structurés différemment avec son feuillet interne et le contenu en eau spécifiques pour chaque liposome.5.2 Hypothèses sur l'évolution moléculaire du processus prébiotique au métabolisme biotique
5.2.1 Introduction
- Il faut bien préciser encore que nous partons d'un milieu réactionnel aux nombreuses sortes de petites molécules, constituant un liquide en équilibre thermodynamique et entouré d'une surface ionisée; et non d'un gel colloïdal concentré en macromolécules arborant de vastes champs électro-dynamiques liés à leur grande surface portant de nombreuses charges positives et négatives à la fois.
Le point de départ du processus prébiotique de la poche de pétrole abiotique est un mélange liquide en contact avec la paroi ionisée négativement des têtes carboxyliques des ag, neutralisée par un mélange de cations. Cet ensemble doit évoluer vers l'état d'une cellule composée d'un gel colloïdal entouré d'une membrane ionisée négativement et neutralisée par quasiment que des cations K+. Il est intéressant de noter ici que les découvertes, dans les recherches sur les origines de la vie, faites sur le piégeage des molécules organiques (dont les P) par les surfaces minérales, mettent en jeu des surfaces ionisées positivement et neutralisées par des anions Cl-, la plus part des cas.
Dans ce qui suit j'ai privilégié le processus de séquestration des aa et d'autres molécules en plus de la fixation des têtes hydrophiles. Mais il n'y a pas que ce processus qui puisse sélectionner les aa dans l'évolution moléculaire. La séquestration apparait comme un tri grossier portant sur le rapprochement des 2 fonctions acide et amine de la tête de l'aa, puis sur leur taille limite. Lorsque j'avais émis l'hypothèse de la séquestration des aa, j'avais prolongé ma réflexion pour imaginer les potentialités des aa et certains ont soulevé des problèmes intéressant vis à vis de cette hypothèse.
Il est hors de question de ne soumettre la sélection des aa qu'au seul processus de la séquestration tant les caractéristiques pour la sélection de 20 aa sont nombreuses. On peut se poser ainsi un grand nombre de questions pour savoir pourquoi ce nombre quasi immuable et pourquoi ceux-là spécialement.- J'ai recencé 5 processus qui peuvent déterminer la forme des aa et la fonction chimique de leurs radicaux:
- Diffusion à travers la membrane (voir chimio-osmose prébiotique)
- Séquestration par la cohésion mécanique du liposome
- Acylation des aa aux phosphates des membranes pour l'encrage des protéines à ces membranes.
- Les contraintes des voies de synthèse métaboliques,synthèses qui peuvent se dérouler aussi dans l'eau loin de la paroi.
- Les fonctions chimiques à effectuer, notamment :
- - Emboitement dans la partie hydrophobe de la bicouche lipidique (aa hydrophobes) et en général participation à la structure 3 D des protéines en intéraction avec d'autres protéines et les membranes par liaisons hydrogènes et attraction électrique entre les dipôles électriques (aa polaires et ionisables);
- - Action à distance par les champs électromagnétiques (aa aromatiques) et
- - Participations aux réactions chimiques proprement dites, c'est-à-dire création et suppression des liaisons covalentes (aa réactifs).
Dans ce qui suit nous allons:
- Illustrer la stratégie mise en oeuvre par le vivant pour élaborer les têtes hydrophiles: Au-delà de la chiralité il y a la conformation spatiale en générale, ce qui explique l'intervention des nucléotides CTP et dCTP pour la mise en place du bras mobile.
- Etudier le cas de la chiralité et de l'homochiralité des sucres dont les origines seraient dans la cohésion mécanique des liposomes: l'hydrogénation du L-glycéraldéhyde-3P en Glycérol-3P sur le bras fixe descrimine le L-glycéraldéhyde en faveur du D-glycéraldéhyde.
- Et mettre en oeuvre la cohésion mécanique du liposome pour l'initialisation du métabolisme prébiotique: Ici nous étudierons l'initialisation et les étapes de l'évolution du métabolisme prébiotique en considérant en plus la séquestration des aa, de la dCTP et de 3 coenzymes. Ces coenzymes interviendront dans l'ordre suivant: aa prébiotiques, B6, dCTP, B1, CTP et THF.
5.2.2 Au-delà de la chiralité il y a la conformation spatiale en générale
- Pour la notation des enzymes du type (EC 4.3.1.7; 260-1-0), voir figure 1 et 2 ci-dessous.
- Dans les chapitres qui suivent j'ai ajouté les abréviations suivantes, parce que répétitives:
- GA, DGA, LGA pour glycéraldéhyde racémique, D et L
- DHA pour dihydroxyacétone;
- PE, PC, PS pour phosphatidyléthanolamine, phosphatidylcholine, phosphatidylsérine.
- PtdGro, Ptd2Gro pour phosphatidylglycérol et diphosphatidylglycérol (ou cardiolipine).
- Pour rechercher les enzymes dans KEGG cliquez ici: [1]
- La chiralité du glycérol-P est établie pendant:
- - l'hydrogénation de la glycérone-P, molécule achirale favorisant la tautomérie;
- - la phosphorylation du glycérol, molécule achirale par symétrie, permettant de fixer le phosphate après la mise en place du glycérol.
- La fixation de la sérine permet de positionner son NH2, comme on l'a vu dans le dessin 1-RG. Elle constitue le seul bras mobile portant une fonction très réactive, la fonction carboxylique.
- La fixation du glycérol en tant que bras mobile, illustre admirablement la nécessité de bypasser la sélection de la conformation adéquate parmi une multitude d'autres conformations. Cette stratégie est utilisée aussi pour la création du bras éthanolamine après fixation de la sérine, comme nous allons le voir dans ce qui suit.
- La création de la PE ne passe pas par la fixation de l'éthanolamine libre. En plus du positionnement de NH2 par la sérine, la PE illustre bien l'importance de la conformation spatiale en général par le fait que l'éthanolamine n'a pas de carbone assymétrique, mais aussi par la nécessité d'éliminer toute entrave à la cohésion mécanique globale du liposome. Parce que l'éthanolamine libre, une molécule très petite, peut échapper au contrôle des macromolécules et mal se positionner ou même établir des liaisons hydrogène avec les acides gras détruisant la cohésion globale.
- Le liposome, en effet, n'est pas rigide. Toutes les molécules PLD de la bicouche sont en mouvement perpétuel tout en assurant la cohésion de l'ensemble. Chez les procaryotes, en effet, l'éthanolamine n'intervient que dans une seule réaction (EC 4.3.1.7; 260-1-0), celle qui la détruit en acétaldéhyde et en NH3. Chez les eucaryotes elle est balisée par une plus grosse molécule qu'est le CDP (EC 2.7.1.82; 2.7.7.14; ) . Ce qui l'empêche de devenir un bras mobile mal emboité. Chez les eucaryotes, autres que les champignons (EC 2.7.8.8; 860-70), la PS est synthétisée à partir de la PE et la sérine (EC 2.7.8.29;0-0-40 ).
- Il est remarquable de noter que la choline libre, avec sa grosse tête de triméthylamine (EC 2.7.8.24; 100-0-0), est utilisée directement sans balise pour synthétiser la PC chez de nombreux procaryotes mais chez aucun eucaryote, comme la sérine le fait avec la PS chez tous les procaryotes et les champignons mais pas chez les eucaryotes supérieurs. Par ailleurs la choline se différencie encore de l'éthanolamine par le fait qu'elle n'est pas détruite immédiatement, mais qu'elle est dégradée en plusieurs étapes en glycine.
- Mais chez tous les êtres vivants la synthèse du bras mobile passe par une estérification intermédiaire inattendue par la CTP. C'est une différence essentielle avec la théorie du métabolisme prébiotique basée sur l'estérification directe des petites molécules. Et cette étape intermédiaire regroupe tous les fondamentaux du métabolisme biotique à savoir un nucléotide (CTP), un désoxynuléotide (dCTP) avec un rapport CTP/dCTP égale à 0.88 (dans Biochemistry of lipids, lipoproteins and membranes 2008, page 74 [7]), un phospholipide et bien sûr une enzyme. Mais ne fait pas intervenir de coenzymes, ni CoA ni B6. Nous avons là réunis en monomères, l'ADN, l'ARN, une protéine et la membrane. Nous allons voir dans ce qui suit que cette étape d'estérification a une seule raison d'être, c'est de mettre en place correctement le bras mobile pour qu'il puisse remplir sa fonction principale, la cohésion mécanique du liposome.

Figure 1. Ce diagramme représente la compilation des organismes possédant un gène donné de la voie métabolique des glycérophospholipides: D'après une impression d'écran de la base de données des voies métaboliques de KEGG.
Les rectangles colorés correspondent à des enzymes membranaires. La compilation est un comptage personnel et approximatif des organismes listés dans la base et consiste en 2 nombres, le 1er celui des procaryotes et le second aux eucaryotes.
Quand l'effectif est faible un 2ème comptage des listes des bases Brenda ou RefSeq (à partir des liens indiqués par KEGG) est accolé à celui issu de la liste de KEGG. Exemple: 0.5.60 correspond à 0 procaryotes dans la liste de KEGG, 5 procaryotes dans Brenda et RefSeq, et 60 eucaryotes dans KEGG. Les dessins des flèches et les noms des molécules sont ceux de KEGG.Figure 2. Analogue à la figure1, d'après les voies métaboliques de KEGG. Ici c'est un montage d'après plusieurs voies métaboliques pour représenter la circulation entre la Sérine, la Glycérone, les acides gras et les phospholipides : Fatty acid Biosynthesis, Fatty acid Metabolism, Glycerolipid Metabolism, Glycerophospholipid Metabolism et Glycolysis.
5.2.3 La cohésion mécanique du liposome est à l'origine de la chiralité et de l'homochiralité des sucres
Dans le métabolisme biotique tous les sucres ont la configuration D pour le 1er carbone asymétrique qui suit la fonction aldéhyde ou cétonique. C'est le cas notamment du D-Glycéraldéhyde à la base de la synthèse de tous les sucres biotiques. Or dans la théorie de la cohésion mécanique du liposome nous n'avons démontré que l'homochiralité du glycérol-3P, de la sérine et éventuellement des autres acides aminés si leur séquestration par les liposomes est effective. En plus le DGA ne fait pas partie de la bicouche lipidique et n'intervient qu'en second lieu à la synthèse du glycérol-3P.
Comment alors s'est établi l'homochiralité du DGA? Peut-on faire intervenir la cohésion mécanique du liposome? Si c'était le cas, il faudrait que le glycéraldéhyde soit séquestré comme les aa, c'est-à-dire prendre la place du bras mobile de la tête hydrophile et qu'il soit de chiralité L. C'est donc le LGA qui aurait pu être séquestré et avantagé par rapport au DGA. Comment alors le LGA a-t-il pu disparaître complètement dans l'évolution moléculaire pour laisser place au seul DGA? Une fois fixé le LGA peut-il être hydrogéné en glycérol? Mais il faut alors supposer une hydrogénation différente de celle du bras fixe.
- La disparition du LGA du métabolisme biotique est due à sa fixation sur le bras fixe en compétition avec la DHA.
- Nous avons vu au chapitre chiralité du glycérol-P que pour les bactéries le glycérol-P qui fixe les acides gras est un L-glycérol-3P. Il a la même conformation que le LGA-3P avec un aldéhyde à la place de l'alcool terminal. Donc le LGA peut prendre la place de la DHA. Ensuite il y a hydrogénation pour obtenir du glycérol-3P, avec la tautomérie du LGA (comme on l'a supposée pour la glycérone) pour faciliter et permettre, dans un premier temps, la configuration spatiale adéquate.
- Du coup l'élimination du LGA entraîne, sinon sa disparition totale (suivant le degré de réversibilité de l'isomérisation prébiotique L <--> D du GA), du moins son désavantage sélectif par rapport au DGA qui restera libre et non fixé aux PLD. L'homochiralité des sucres peut alors commencer. Nous voyons ainsi que l'homochiralité des aa provient de leur séquestration ou de leur fixation (sérine) dans les bras libres des PLD et se fait encore dans le métabolisme biotique, alors que l'homochiralité des sucres a dû se faire une fois seulement pendant le métabolisme prébiotique, le LGA disparaissant après complètement.
- Chiralité du PtdGro
- Le glycérol-P du bras mobile de PtdGro est le L-glycérol-3P. En effet il a même configuration que la L-sérine avec l'alcool terminal (CH2OH) à l'extérieur de la membrane. C'est le même que celui du bras fixe puisque celui-ci a l'alcool terminal disposé vers l'intérieur de la membrane (paragraphe chiralité du glycérol-P). PtdGro est décliné, dans Kegg, en phosphatidylglycérol sans préciser la chiralité du glycérol. A mon avis il faut le nommer phosphatidyl-L-glycérol (sans ajouter 3P qui porterait à confusion). D'ailleurs la réaction est écrite clairement dans Kegg [9]:
- CDP-diacylglycerol + sn-glycerol 3-phosphate = CMP + 3(3-sn-phosphatidyl)-sn-glycerol 1-phosphate (EC 2.7.8.5; 1200-110)
- le P de sn-glycérol 1-P se réfère au P terminal, alors que le substrat est bien sn-glycérol 3-P.
- Le PtdGro existe aussi dans les têtes hydrophiles des Archées [10]. On a vu que chez les bactéries que le L-glycérol-3P pouvait venir de l'hydrogénation de DHA ou du LGA-3P. Si pour les archées le D-glycérol-3P devait provenir, au début de l'évolution prébiotique, aussi du DGA-3P on aurait eu une homochiralité L de tous les sucres. Ce qui n'est pas le cas. Donc les archées, ayant tous leurs sucres de chiralité D, sont apparus après les bactéries.
- Il est à noter cependant que la synthèse du D-glycérol-3P et son utilisation restent toujours possibles même dans le métabolisme biotique des bactéries. En effet le Ptd2Gro (EC 278.-, Cls;700-100), ubiquitaire, a un glycérol flanqué de 2 acides phosphatidiques avec 4 liaison esters hydrolysables. La libération du glycérol-P peut se faire de 2 manières possibles donant chacune un glycérol-3P ou un glycérol-1P (dans Biochemistry of lipids, lipoproteins and membranes 2008, page 75) [8]).
- On aurait tendance à dire que les premiers bras mobiles qui seraient apparus, aient été des L-glycérol-3P puisque ceux-ci ont été déjà synthétisés à la première étape de la constitution de la tête hydrophile. Mais il faut remarquer que la synthèse du PtdGro demande une étape de plus que celle du PS, à partir d'un glycérol qu'il faut détacher du bras fixe; que le glycérol risque, s'il était l'unique bras mobile, d'établir dans certaines conditions de température et de pression des liaisons hydrogènes généralisées, puisqu'il n'est pas ionisable, ce qui cristallisera le liposome.On voit alors l'importance de la sérine qui peut être synthétisée abiotiquement [5], même en petite quantité, ou bien qu'elle puisse être synthétisée en une seule réaction enzymatique (en présence de B6), dans le métabolisme biotique (EC 431.17; 800-20, et 431.19; 930-100). La sérine est intéressante parce que c'est un acide aminé réactif par sa tête zwitterionique et qu'elle peut en une seule étape donner PE dont l'amine est plus réactive, encore, que le glycérol.
5.2.4 La CMP positionne le tétraèdre du phosphate pour estérifier le bras libre dans la bonne configuration nécessaire à la cohésion mécanique du liposome.
- Nous avons vu au paragraphe sur la conformation spatiale en générale, la différence essentielle et inattendue entre estérifications prébiotiques hypothétiques et les estérifications biotiques. En considérant la configuration spatiale en général au lieu de la chiralité seule, il apparaît clairement qu'en prébiotique, le nombre de configurations explose lorsqu'on tient compte de la rotation du phosphate libre avant son estérification par la sérine, le glycérol, l'éthanolamine ou la choline.
- Dans la fixation du glycérol du bras libre, en métabolisme biotique, le phosphate (du glycérol-3P) a servi à le baliser et peut-être à forcer sa fixation dans la position adéquate de la cohésion mécanique. Mais le P du PtdGro-P alourdit le bras mobile (EC 2.7.8.5; 1200-110). Le phosphate est expulsé rapidement dans la réaction EC 3.1.3.27 (500-4-0) chez la plupart des procaryotes.
- Il y a 3 enzymes qui positionnent le phosphate, (Ec 2.7.7.41; 1200-140) pour la sérine et le glycérol, (Ec 2.7.7.14; 0-0-20) pour l'éthanolamine et (Ec 2.7.7.15; 25-70) pour la choline. Nous voyons que seule la première est généralisée à tous les êtres vivants, alors que les 2 autres sont plutôt propres aux eucaryotes seulement.
-
5.2.5 La cohésion mécanique du liposome et l'initialisation du métabolisme prébiotique
Remettons-nous dans les conditions de la poche de pétrole prébiotique comme on l'a décrit dans les conditions de la poche de pétrole prébiotique, et reconsidérons l'initialisation du métabolisme prébiotique à la lumière des résultats obtenus jusqu'ici dans une vésicule aqueuse de la phase huile.
Nous avions admis que l'initialisation du métabolisme ne pouvait démarrer qu'à conditions que les molécules nécessaires puissent être piégées par la paroi de la vésicule, déplaçant régulièrement les équilibres thermodynamiques du contenu aqueux. Excéptée l'hydrogénation du DHA ou du GA par H2, hydrogénation dont on avait supposé l'existence, toutes les autres réactions de la synthèse des têtes hydrophiles sont des estérifications très lentes si elles se font sans enzymes. Ce sont dans l'ordre: - - Phosphorylation du DHA et du GA;
- - Estérification de ces molécules phosphorylées aux ag après hydrogénation in situ, catalysé par le phosphate et la surface carboxylique.
- - Fixation de l'éthanolamine ou de la sérine ou du glycérol. En supposant que ce dernier puisse exister, par exemple par son hydrolyse de la paroi.
- Les résultats de la réflexion sur la théorie de la cohesion mécanique du liposome montre que celle-ci crée un cercle vertueux: l'augmentation du nombre de têtes hydrophiles de même chiralité augmente la cohésion de la vésicule qui favorise à son tour la fixation de molécules de même chiralité, formant de nouvelles têtes hydrophiles.
- Ce cercle vertueux de la cohésion mécanique va aussi créer un autre cercle vertueux basé sur la catalyse: la L-sérine des têtes hydrophiles a son carboxyle libre et très réactif; la cohésion mécanique par ailleurs concentre de plus en plus de L-aa sur une surface de plus en plus en grande de têtes hydrophiles, ce qui favorise le regroupement d'aa en entités catalytiques de plus en plus efficaces.
- Certaines études (Wieczorek, 2009 [8]) ont montré par exemple que le dipeptide Ser-His, attaché à la paroi, est très réactif et est à l'origine de nombreux processus qui se déroulent dans le liposome. On le trouve dans les sites actifs de nombreuses enzymes. Dans le cas du métabolisme prébiotique où les réactions sont très lentes, une seule molécule de ce dipeptide peut avoir une forte accélération de l'évolution moléculaire.
- Séquestration de certains coenzymes.
- Le processus de séquestration devient réellement intéressant avec la séquestration de précurseurs de quelques coenzymes (voir leurs dessins ci-dessous) comme la pyridoxamine pour le pyridoxal-P, le noyau pyrimidinique de la thiamine, le noyau pteridinique de l'acide folique et la désoxycytidine. Ce sont de petites molécules de la taille des aa avec une amine et un hydroxyl comme l'éthanolamine. En plus la désoxycytidine se fixe carrément au PLD, sous forme dCMP, comme la sérine dans le métabolisme biotique.
- L'analogie des 3 autres coenzymes avec la dCTP est encore plus frappante puisque leurs intermédiaires sont phosphorylés avant d'être convertis en leur forme active, comme la desoxycytidine: c'est ainsi que le noyau pyrimidinique de la thiamine (EC 271.49, 2747) et le noyau pteridinique du folate (EC 2763) sont phosphorylés avec 2 phosphates dans le métabolisme biotique. Curieusement la pyridoxamine, quand a elle, est convertie en la forme active, le pyridoxal-5P chez Clostridium kainantoi par une transaminase (EC 261.54) à partir du D-Ala ou du D-Glu. Une transaminase de ce type se fait normalement avec un L-aa et a comme coenzyme le pyridoxal-5P.
- Nous allons supposé que dans le métabolisme prébiotique, ces précurseurs sont d'abord séquestrés (ou fixés?) sur la paroi avant d'être convertis en leur forme active. Notamment nous allons voir (tableau 1 étape 5) que la dCTP primordiale pour le métabolisme biotique pourrait être intéressante de l'introduire avant l'apparition de la CTP puisque celle-ci nécessite pour sa synthèse l'ATP (étape 6), alors que la dCTP pourrait apparaitre avant (étape 4).
- Table 1. Coenzymes prébiotiques: Les dessins et les codes de ces molécules sont ceux de la base de données KEGG.


-thiazole")
- Thiamine 4-Amino-5-hydroxymethyl-2-methylpyrimidine 4-Methyl-5-(2-hydroxyethyl)-thiazole




- Acide folique 2-Amino-4-hydroxy-6-hydroxymethyl-7,8-dihydropteridine Acide 4-Aminobenzoïque Glutamate
-



- Desoxy-cytidine Cytosine 2-Desoxy-D-ribose


-
Pyridoxamine Pyridoxal
- Les étapes de l'initialisation du métabolisme prébiotique, Tableau 1:
Nous allons maintenant dérouler le scénario de l'initialisation du métabolisme prébiotique en commençant par la synthèse des têtes hydrophiles générant la cohésion mécanique et la différenciation entre la paroi riche en L-aa séquestrés et l'intérieur liquide riche en D-aa.
Puis interviendrait la séquestration de B6 qui jouerait le rôle de coenzyme pour les groupes de L-aa de la paroi. Je fais intervenir B6 en 1er par analogie, parce que dans le métabolisme biotique, elle active d'innombrable réactions entre les aa plus NH3. Décuplant ainsi le cercle vertueux de la catalyse.Tableau 1. La cohésion mécanique et les étapes de l'initialisation du métabolisme.Fixation Séquestre Produits Cumul Disparition Utilisation aa présents Commentaires à partir du site KEGG (analogie des groupes de L-aa séquestrés avec les enzymes). 0 Vésicule. Energétique prébiotique : Formose. Synthèse hydrothermale des aa : ADEGS. (pour l'analogie avec la synthèse des L-aa voir le site KEGG). 1 DHA LGA L-ser Têtes hydrophiles PS DGA D-ser cohésion LGA H2 ADEGS Hydrogénation catalytique prébiotique, non enzymatique, de DHA et LGA et non du DGA. Accumulation de D-ser et l'homochiralité des sucres est due à la fixation du LGA. 2 L-aa groupes L-aa D-aa L-ser libre initial ADEGS ADEGS Différenciation entre surface (L-aa) et intérieur aqueux (D-aa). 3 B6 L-ser
NQCTWY ADEG
PE PtdGro Ptd2Grocohésion
2-oxo-acidesaa libres initiaux H2S NH3 indol phenol acetaldéhyde NQCTWY ADEGS B6 et le regroupement des L-aa homogènes catalysent mieux les réactions à 1 étape sans ATP, utilisant NH3. Les interconversions entre les aa accumulent les oxo-acides qui avec DHA et DGA ils préparent le métabolisme intermédiare des acides carboxyliques. 4 dR-1P dATP DGA-3P+acetal Adenine Réactions sans coenzymes : 4124, 5427 donnent le D-dRibose-1P. Puis avec Adénine : 2421,271.76,2743,2746 donnent dATP. 5 CTP cytosine cytosine+dR-1P Cohésion Cytosine dR-1P dATP L'équivalent de 2421 n'existe pas pour la cytosine ; les groupes de L-aa pourraient catalyser la synthèse de dCTP sur place. 6 B1 ATP CTP NAD B6 SAM FAD FMN Biotine FHKPIVM
PCCohésion H2 initial B6 initial Bases nicotinate DHA+DGA dATP FHKPIVM NQCTWY ADEGS B1 est constituée de m1 et m2, dont m1 peut être séquestrée. La synthèse de B1 peut se faire sur place comme pour dCTP. Avec B1, 412.13, 313.11, 2211, 5131, 5316 et à partir de DHA+DGA on obtient R-5P qui avec dATP donne PRPP (2761) et R-1P (5427).
A+R-1P+PRPP+PPP → ATP : 2421, 2428, 2743, 2741.
C+R-1P+ATP → CTP : 2422, 271.48, 274.14, 2746.
N+PRPP+ATP+NH3 → NAD : 242.11, 2771, 6351.
D-Ribulose-5P (5131)+DGA-3P+L-gln → Pyridoxal-P (B6) : 4.-.-.-, YaaD, Pyridoxal biosynthesis lyase pdxS. 2.6.-.-, YaaE, Glutamine amidotransferase subunit pdxT.7 THF CoA acides gras LR DHA DGA de formose Phosphate LR FHKPIVM NQCTWY ADEGS THF est constitué de m3 et m4 qui peuvent être séquestrées.
Pyruvate+B1+NAD+THF+ATP+L-asp+L-cys → CoA : 2216, 11.86, 4219, 212.11, 111.169, 411.11, 6321, 271.33, 6325, 411.36, 2773, 271.24.. 8 Formation des liposomes : passage à l'énergétique membranaire. . 9 DHA DGA Bases nicotinate indol phenol Bases nicotinate indol phenol initiaux Metabolisme actuel L-aa actuels Voir site KEGG 10 B1 THF B1 THF initiaux Metabolisme actuel L-aa actuels Voir site KEGG - Etape 0.
- - H2 H2S CO2, N2 puis NH3 les gaz des évents hydrothermaux (Charlou [9] 2002, Proskurowski [10] 2008 ).
- - Phosphates et polyphosphates des fonds marins (Arrhenius 1997 [11]).
- - alcanes, acides gras, alcools et aldéhydes du processus Fischer-Tropsch, dont l'acétaldéhyde (Rushdi[12] 2001, McCollom 1999,[13] 2006 [14]).
- - DHA GA glyoxal par hydroformylation ou formose à 120 bars et 140 °C [3].
- - ADEGS aa obtenus dans les expériences hydrothermales avec des molécules azotées [4].
- - Précurseurs des coenzymes et noyau aromatiques en très petites quantités de la poche prébiotique (hypothèse):
- Bases nucléiques, A G C U T, pyridoxamine, les 2 noyaux de la thiamine, les 2 noyaux du folate et le nicotinate pour le NAD.
- Etape 1.
- Synthèse de la tête hydrophyle suivant l'analyse des paragraphes précédent. Fixation sur la paroi de L-ser, DHA et LGA, accumulation de DGA et D-ser dans l'eau. Origine de l'homochiralité des sucres via DGA. Cercle vertueux synthèse_des_têtes/cohésion_mécanique.
- Etape 2.
- - Séquestration des L-aa par les parois et concentration des D-aa dans l'eau. L-ser disparait petit à petit de l'eau avec la diminution de la synthèse hydrothermale des aa avec le temps.
- - Regroupement des L-aa sur la paroi pour constituer des pseudo-enzymes. Elles ne sont peut-être pas très efficaces, mais de multiples combinaisons plus ou moins éphémères sont possibles. Plus la surface des têtes hydrophiles grandit plus les groupes pourraient être solides et nombreux, plus ils seront coopératifs dans la catalyse et leur regroupement même. C'est le cercle vertueux de la catalyse soutendu par la cohésion mécanique.
- Etape 3.
- - L'accumulation de DGA et de D-ser déplacent très lentement les équilibres vers L-ser.
- - La séquestration de B6 accélère l'isomérisation de D-ser en L-ser et la désamination de D-ser en pyruvate (Ec 431.18) puis son amination en L-ser (EC 431.17).
- - Sous l'action de B6, synthèse de nouveaux aa: à partir du pyruvate, du NH3 et de l'indole (Trp) ou du phénol (Tyr); avec la thiolyse par H2S des têtes hydrophiles (Cys); avec la condensation de la Gly et de l'acétaldéhyde en Thr; et l'amination du Glu en Gln et Asp en Asn (voir KEGG pour l'analogie avec les enzymes).
- - Sous l'action de B6, décarboxylation de la sérine de PS pour donner PE (EC 411.65). L'éthanolamine très peu produite dans la synthèse hydrothermale des aa, a pu être fixée à la place de la sérine mais beaucoup plus difficilement comme on l'a vu précédemment. Il y a renforcement de la cohésion mécanique par la PE car elle n'a pas de tête réactive.
- - Sous l'action de B6, désamination de Thr pour donner le 2-oxo-butanoate (EC 4125) nécessaire plus loin pour la synthèse de Val Leu Ile.
- - Transaminations entre les aa (ADEGS plus les nouveaux aa, CNQTWY) pour la production de L-ser, ce qui accumule les 2-oxo-acides.
- - DGA, DHA, 2-oxo-acides, NH3, H2S et les aa ADEGS+CNQTWY constituent le métabolisme intermédiaire prébiotique.
- - Tous ces nouveaux produits grâce à B6 s'obtiennent en une seule réaction.
- Etape 4.
- - Synthèse sans coenzyme du premier desoxy-pentose en 2 réactions (EC 4124, 5427 [15]) dont la 2ème est auto-catalytique (formation de ribose biphosphate): DGA-3P + acétaldéhyde = D-dRibose-1P.
- - Synthèse de la désoxy-Adénosine sans coenzyme (EC 2421), puis 2 phosphorylations avec P et PPP donnent la dADP (EC 3135, 2743).
- - Le passage à la dATP nécessite l'ATP dans le métabolisme biotique (EC 2746). Je suppose qu'en prébiotique qu'il est tout à fait possible que la dATP se forme très lentement par auto-catalyse (en confondant dATP et ATP dans EC 2746) ou en présence de polyphosphates comme les 2 réactions qui la précèdent. D'ailleurs c'est une estérification et j'avais basé l'évolution moléculaire prébiotique sur les estérifications (voir paragraphe 5.1.2).
- Etape 5.
- - Cette étape peut paraitre théoriquement superflue, mais l'étape 6 qui suit nécessite la thiamine et 7 réactions pour arriver au Ribose-1P. Or nous avons vu à la séquestration de la thiamine, du folate et de la dCTP qu'il fallait une condensation in-situ des 2 parties pour former chaque coenzyme. La dCTP devrait paraitre bien avant la CTP.
- - Dans le métabolisme biotique, il n'existe pas d'équivalent pour la dCTP de la EC 2421 de l'étape 4. Or le passage par la dCTP ou la CTP pour la fixation du bras mobile (EC 277.41), quelque soit le bras, semble primordiale. Aussi la formation in-situ en présence de groupement de L-aa séquestrés apporterait rapidement une grande cohésion mécanique. La dCTP se comporte comme un coenzyme puisque la dCTP, après hydrolyse peut être regénérée par phosphorylation de la dCMP.
- Etape 6.
- - Synthèse in-situ de B1 (voir le dessin des molécules ci-dessous).
- - C'est l'accumulation de DGA dès la première étape du scénario qui va favoriser, par les déplacements des équilibres thermodynamiques, sa condensation avec la DHA, toutes les 2 sous forme phosphorylée: DGA-3P + DHA-P (EC 412.13).
- - ATP: Il faut 6 réactions et la participation de dATP et de B1, seules coenzymes, pour arriver à la molécule centrale PRPP puis une isomérisation pour arriver au Ribose-1P nécessaire à la synthèse de l'ATP à partir de l'Adénine et de PPP.
- - La synthèse de la CTP nécessite l'ATP.
- - Le NAD nécessite le nicotinate et l'ATP.
- - La synthèse de la S-adénosyl-méthionine (SAM), des flavines (FAD et FMN) et de la biotine ne nécessite que les coenzymes prébiotiques déjà créées.
- - Le Pyridoxal-5P, B6, part d'un intermédiaire de la synthèse du D-Ribose-1P, le D-Ribulose-5P: D-Ribulose-5P+DGA-3P+L-Gln (EC 5131).
- - La cohésion va se développer à grande vitesse avec la CTP; remplacement de l'hydrogénation prébiotique (H2) par l'hydrogénation par le NAD et le B6 est synthétisé de novo.
- - 7 nouveaux aa peuvent être synthétisés dont l'histidine qu'on a vu, importante pour les sites actifs des protéines: FHKPIVM.
- - La synthèse de la méthionine permet la synthèse de SAM et de là la production de PC.
- Etape 7.
- - Synthèse in-situ du folate (THF) (voir le dessin des molécules ci-dessous).
- - Synthèse du coenzyme A, CoA et des 2 derniers aa, LR, synthèse des ag.
- Etape 8.
- - La formation du liposome avec le passage de la vésicule dans la phase eau, comme on l'a vu est nécessaire parce qu'à un moment ou un autre, l'énergie prébiotique limitée à la vésicule (formose: DHA et DGA), s'épuisera. L'énergie prébiotique sera remplacée par l'énergétique membranaire qui suit l'évolution moléculaire supposée dans l'article de la chimio-osmose prébiotique.
- - Les molécules de base du métabolisme, notamment le phosphate, arrivent alors par diffusion d'abord, par flip-flop physique pour le phosphate, puis par les porines primitives équivalentes aux ionophores et enfin par les transporteurs protéiques.
- - Les complexes énergétiques protéiques s'installent dans la membrane par migration des aa de l'intérieur ou par diffusion de ceux venant de l'extérieur.
- Etape 9.
- - Avec le métabolisme membranaire, synthèse de DHA et DGA de novo, des acides gras et des coenzymes membranaires comme les cytochromes. Remplacement des noyaux aromatiques: indol, phénol, nicotinate et les bases nucléiques.
- Etape 10.
- - Remplacement de la Thiamine et du folate (THF).
6. Références
- ↑ Dorota Wolicka, Andrzej Borkowski, and Dariusz Dobrzynski: Interactions between Microorganisms, Crude Oil and Formation Waters. Geomicrobiology Journal, 27:43–52, 2010.
- ↑ Tupi
- ↑ 3,0 et 3,1 Zoltán Györgydeák,István F. Pelyvás. Monosaccharide sugars: chemical synthesis by chain elongation, degradation...(Page 8). Academic Press 1998.
- ↑ 4,0 , 4,1 et 4,2 A. D. Aubrey & H. J. Cleaves & Jeffrey L. Bada: The Role of Submarine Hydrothermal Systems in the Synthesis of Amino Acids. Orig Life Evol Biosph (2009) 39:91–108 DOI 10.1007/s11084-008-9153-2
- ↑ Ifremer fiche 8
- ↑ Daniel Segré and Doron Lancet: Composing life. EMBO Reports vol.1 no.3, pp 217–222, 2000. Segre_Lancet_EMBOrep_2000.pdf
- ↑ 7,0 et 7,1 Biochemistry of lipids, lipoproteins and membranes 5th edition, 2008: edited by D.E. Vance and J.E. Vance; Elsevier
- ↑ Gorlero M, Wieczorek R, Adamala K, Giorgi A, Schininà ME, Stano P, Luisi PL. (2009) Ser-His catalyses the formation of peptides and PNAs. FEBS Lett. 583(1):153-6.
- ↑ Charlou J.L., Donval J.P., Fouquet Y., Jean-Baptiste P., Holm N., « Geochemistry of high H2 and CH4 vent fluids issuing from ultramafic rocks at the Rainbow hydrothermal field », Chemical Geology, vol. 191, 2002, p. 345-359. sciencedirect
- ↑ Giora Proskurowski, Marvin D. Lilley, Jeffery S. Seewald, Gretchen L. Früh-Green, Eric J. Olson,1 John E. Lupton, Sean P. Sylva, Deborah S. Kelley: Abiogenic Hydrocarbon Production at Lost City Hydrothermal Field . Science vol 319, 1 février 2008 sciencemag
- ↑ G. Arrhenius, B. Sales, S. Mojzsis and T. Lee : Entropy and Charge in Molecular Evolution-the Case of Phosphate Journal of Theoretical Biology Volume 187, Issue 4, 21 August 1997, Pages 503-522 sciencedirect
- ↑ Ahmed I. Rushdi, and Bernd R.T. Simoneit: Lipid formation by aqueous fischer-tropsch-type synthesis over a temperature range of 100 to 400 °C. Origins of Life and Evolution of Biospheres (2001) 31: 103–118.
- ↑ T.M.McCollom et al. 1999: Lipid synthesis under hydrothermal conditions by fischer-tropsch-type reactions . Origins of Life and Evolution of the Biosphere 29: 153–166, 1999
- ↑ T.M.McCollom et al. 2006:Carbon isotope composition of organic compounds produced by abiotic synthesis under hydrothermal conditions. Earth and Planetary Science Letters Volume 243, Issues 1-2, 15 March 2006, Pages 74-84
- ↑ http://www.genome.jp/kegg/pathway/map/map00030.html
1 commentaire Suivre le flux RSS des articles de cette rubrique Suivre le flux RSS des commentaires de cette rubrique
Suivre le flux RSS des articles de cette rubrique Suivre le flux RSS des commentaires de cette rubrique



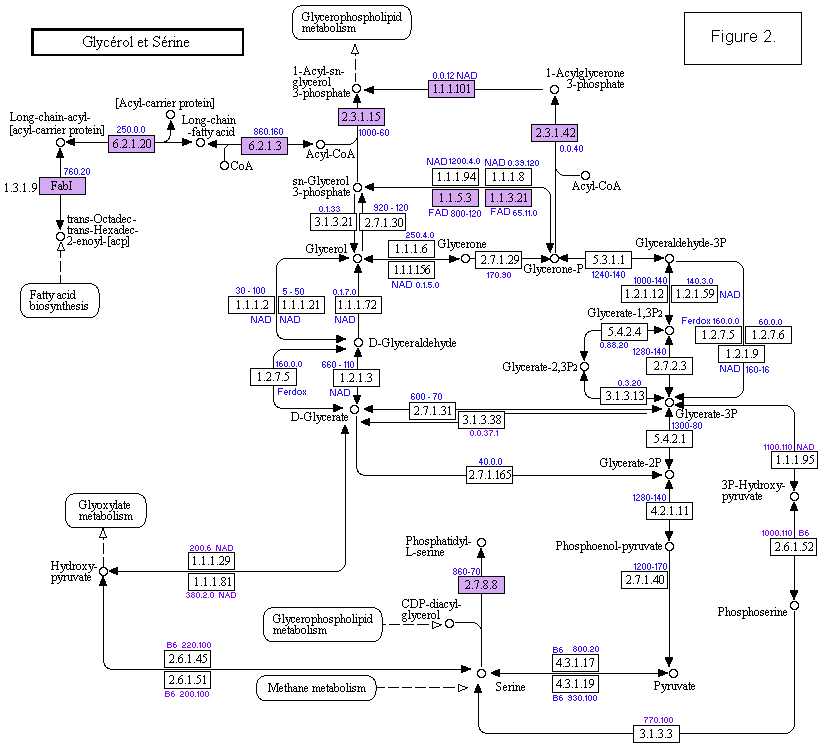
|
|
|
|